Avicenna Journal of Clinical Microbiology and Infection. 12(4):180-187.
doi: 10.34172/ajcmi.3738
Original Article
Molecular Profiling of Antibiotic Resistance and Virulence in Multidrug-Resistant Uropathogenic Klebsiella pneumoniae
Anandha Krishnan Rajaram Heamchandsaravanan Conceptualization, Data curation, Methodology, Resources, Validation, Writing – original draft, Writing – review & editing, 1 
Ishrath Razia Riaz Data curation, Formal analysis, Methodology, Writing – original draft, Writing – review & editing, 1
Maheswari Paulraj Resources, Visualization, 1
Divyapriya Dhayalan Formal analysis, Writing – review & editing, 1
Ragala Venkata Nandha Methodology, 1
Prabu Dhandapani Conceptualization, Investigation, Project administration, Resources, Supervision, Validation, Visualization, 1, *
Author information:
1Department of Microbiology, Dr. ALM PG institute of Basic Medical Sciences, University of Madras, Taramani Campus, Chennai, Tamilnadu, India
Abstract
Background: Urinary tract infections (UTIs) constitute a serious public health concern, with Klebsiella pneumoniae being a prevalent cause worldwide. This bacterium is recognized for elevated recurrence rates and significant antibiotic resistance. The complex virulence mechanisms of K. pneumoniae contribute to its persistence within the host. Nevertheless, it is still unknown how particular virulence genes and antibiotic resistance relate to uropathogenic K. pneumoniae. This study examined the relationship between pathways of virulence and antibiotic resistance in the isolates of uropathogenic K. pneumoniae.
Methods: Overall, 207 uropathogenic K. pneumoniae isolates were analyzed for antibiotic susceptibility. Then, polymerase chain reaction (PCR) was used to identify resistance (blaSHV, blaTEM, blaNDM, blaCTX-M, blaKPC, and blaOXA) and virulence (Ompk35, Ompk36, Mdtk, AcrAB, TolC, mrkD, allS, ybtS, entB, kfu, and iutA) genes. Eventually, enterobacterial repetitive intergenic consensus (ERIC)-PCR was utilized to assess the genetic diversity of the strains.
Results: All multidrug-resistant strains (100%) carried the Mdtk and AcrAB efflux pump genes. Virulence genes (e.g., mrkD and entB) were common across the isolates. Several genes were shown to be prevalent in β-lactam resistance, including blaSHV (95.8%), blaCTX-M (70.8%), blaNDM (62.5%), blaTEM (41.7%), and blaOXA (33%). Finally, ERIC-PCR produced distinct banding profiles between 50 and 1500 base pairs, with each isolate demonstrating between one and eight bands.
Conclusion: Our findings emphasize the connection between K. pneumoniae virulence genotypes and antibiotic resistance. Understanding these associations is crucial for managing and preventing K. pneumoniae-related UTIs effectively.
Keywords: Antimicrobial resistance, ERIC-PCR, K. pneumoniae, Uropathogenic, Virulence genes
Copyright and License Information
© 2025 The Author(s); Published by Hamadan University of Medical Sciences.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (
https://creativecommons.org/licenses/by/4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Please cite this article as follows: Heamchandsaravanan AR, Riaz IR, Paulraj M, Dhayalan D, Nandha RV, Dhandapani P. Molecular profiling of antibiotic resistance and virulence in multidrug-resistant uropathogenic Klebsiella pneumoniae. Avicenna J Clin Microbiol Infect. 2025;12(4):180-187. doi:10.34172/ajcmi.3738
Introduction
Recently, many centres around the world have undergone outbreaks of multidrug-resistant (MDR) Enterobacteriaceae infections, along with Klebsiella pneumoniae, highlighting the urgent need for effective control and treatment strategies on a global scale. The most important pathogen among the genus is K. pneumoniae, which causes a variety of diseases in hospitals, assisted living homes, and communities worldwide. These infections can affect the lungs, urinary tract, abdominal cavity, surgical sites, and soft tissues and can even lead to bacteremia (1). Despite being the second most frequent cause of urinary tract infections (UTIs) after Escherichia coli, K. pneumoniae is more harmful than a related species (2). K. pneumoniae is a major opportunistic pathogen involved in a wide spectrum of infections, including those affecting the respiratory tract, urinary system, bloodstream, and soft tissues, particularly in healthcare environments (3). Approximately 12% of UTIs are attributed to K. pneumoniae, with this number rising rapidly worldwide, particularly in Asia. The spread of bacteria that produce extended-spectrum beta-lactamases (ESBLs) and are resistant to antibiotics is mostly due to this increase (4). Drug-resistant K. pneumoniae is becoming more common, which has increased its therapeutic effect and made treatment more difficult. Most uropathogenic K. pneumoniae isolates have developed resistance mechanisms against various antimicrobial agents, particularly β-lactams, cephalosporins, and polymyxins. Antimicrobial-resistant uropathogenic K. pneumoniae surveillance is essential to enhance infection control measures, particularly in the healthcare sector, and to aid clinicians in the selection of empirical therapy. According to previous research, virulence gene profiling and genotyping may provide some insights into K. pneumoniae infections (5). Siderophores, fimbriae, outer membrane porins, and a protective capsule are important components of the pathogenicity of this pathogen. These features support the growth of drug resistance, the attachment of bacteria to uromucosal surfaces, the initiation of inflammatory reactions, and the spread of disease throughout the urogenital tract (6,7). The bacteria are protected from phagocytosis by the bacterial capsule, which is made up of strain-specific capsular polysaccharides called K antigens. Notably, K1 and K2 are associated with serious human infections out of the 77 capsular serotypes that have been discovered (8). Genes linked to virulence, such as those governing siderophore production (entB and ybtS), adhesins (mrkD and fimH), iron uptake (kfu and iutA), and allantoin metabolism (allS), play an essential role in enabling K. pneumoniae to adhere to and invade host tissues. The harmful effects of K. pneumoniae infections, especially those contracted in hospital settings, are significantly influenced by these genes (9,10). Six virulence genes (mrkD,ycfM,fimH,ybtS,entB, and kfu) and three β-lactamase-encoding genes (blaKPC, blaCTX-M-Gp1, and blaOXA-1-like) were observed in K. pneumoniae in a prior study (11). ESBLs result from the production of enzymes encoded by the blaSHV,blaTEM, and blaCTX -M genes, with nearly 300 variations described (12). Additionally, AcrAB-TolC efflux pumps (mdtk,AcrAB, and TolC) and outer membrane porins (Ompk35 and Ompk36) are crucial for antimicrobial resistance in K. pneumoniae. The presence of these virulence factor genes influences the pathogenicity and genetic diversity of uropathogenic isolates. Prior research has highlighted the prevalence of various genotypes with distinct virulence profiles in MDR uropathogenic K. pneumoniae strains (13). Detailed knowledge of genotype, antibiotic resistance, and virulence pattern of uropathogenic K. pneumoniae is necessary for reducing the risk of outbreaks. Enterobacterial repetitive intergenic consensus-polymerase chain reaction (ERIC-PCR), the amplification of repetitive sequences in the bacterial genome, is a cheaper and faster genotyping method for determining clonal diversity among uropathogenic K. pneumoniae isolates. Differences in the positions and the number of repetitive sequences will result in a different banding pattern by gel electrophoresis (14). Therefore, the current study aims to examine the connection between uropathogenic MDR K. pneumoniae’s genetic diversity and virulence genes.
Materials and Methods
Sample Collection and Study Design
This prospective analytical study was conducted at the Department of Microbiology, Dr. ALM Postgraduate Institute of Basic Medical Sciences, University of Madras, from September 2017 to January 2021. A total of 207 laboratory-archived uropathogenic K. pneumoniae isolates were collected from patients with severe UTIs and kept at -20 °C. Standard biochemical analysis was used to identify and resuscitate the isolates.
Antibiotic Susceptibility Test
The antibiotic susceptibility of K. pneumoniae isolates was evaluated using the Kirby-Bauer disc diffusion method with commercially available antibiotic discs (Himedia, Mumbai, India). Initially, ESBL production was screened using the discs of ceftazidime (30 μg), aztreonam (30 μg), cefotaxime (30 μg), and ceftriaxone (30 μg). Confirmation was performed using the combined disc method with ceftazidime (30 μg) and ceftazidime/clavulanic acid (30/10 μg), considering a ≥ 5 mm zone difference as indicative of ESBL production. In addition, metallo-beta-lactamase (MBL) production was detected in imipenem-resistant strains using the combined disc test with imipenem (10 μg) discs, one supplemented with 10 μL of 0.5 M ethylenediaminetetraacetic acid (pH: 8.0), and a ≥ 7 mm zone difference indicating MBL production. This procedure was similarly applied to meropenem. Moreover, AmpC beta-lactamase production was detected using cefoxitin (30 μg) discs. Fluoroquinolone resistance was confirmed using nalidixic acid (30 μg), ciprofloxacin (5 μg), norfloxacin (10 μg), ofloxacin (5 μg), and levofloxacin (5 μg) discs, while aminoglycoside resistance was tested with gentamicin (10 μg) and amikacin (30 μg) discs. Finally, zone diameters were determined and reported in accordance with the Clinical and Laboratory Standards Institute’s (CLSI’s) recommendations.
Colistin Resistance Screening
For the initial screening of colistin resistance, K. pneumoniae isolates were inoculated in 0.9% sodium chloride and adjusted to 0.5 McFarland standards. Then, eosin methylene blue agar plates containing 3.5 μg/mL colistin were streaked with a 10 μL inoculum. Next, the plates were incubated in an aerobic environment for 24–48 hours at 37 °C. Colistin susceptibility was then determined by the absence of growth or limited growth in the primary streak (susceptible) versus growth in all three streaks (resistant), following CLSI–the European Committee on Antimicrobial Susceptibility Testing (EUCAST) guidelines. Suitable control strains were present. Subsequently, colistin-resistant isolates were subjected to further testing in accordance with CLSI standards (2018) using the broth microdilution method. Minimum inhibitory concentration values were interpreted based on CLSI-EUCAST criteria: ≤ 2 µg/mL as susceptible and > 2 µg/mL as resistant.
Deoxyribonucleic Acid Extraction
Using the boiling lysis method, DNA was extracted based on our previously published research (15). A Nanodrop spectrophotometer (Nanovue Plus; GE Healthcare Life Sciences, United States) was used to measure the amount and quality of the extracted DNA and then stored at -20°C for further use.
Detection of Virulent and Multidrug Resistance Genes
The PCR was utilized to identify capsular serotypes (K1 and K2) and virulence genes (ybtS, mrkD,rmpA,entB,iutA,kfu, and allS) (10), cephalosporin resistance genes (blaTEM,blaSHV, and blaCTX -M), carbapenem resistance genes (blaNDM, blaKPC, and blaOXA) (16), transport system genes (Ompk35, Ompk36,TolC,mdtk, and AcrAB), and mcr genes (mcr1-8) (17). The specific primers employed in this study are presented in Supplementary file 1, and Table 1 provides the PCR programs for different genes. Amplicons were confirmed using 1.5% agarose gel electrophoresis, stained with ethidium bromide, and visualized with a Carestream Gel Logic 212 Pro gel documentation system (USA).
Table 1.
Multiplex PCR Protocols and Conditions Used for Detecting Capsular Serotypes, Virulence Factors, Antibiotic Resistance Genes, and Transport System Genes
|
Target Genes
|
PCRMix(µL)
|
Conditions
|
| Capsular K1 and K2 |
10: 5 PCR master mix, 2 DNA templates, 1 milliQ water, 0.5 of each primer |
95 °C for 15 minutes; 30 cycles of 94 °C for 30 seconds, 60 °C for 90 seconds, and 72 °C for 60 seconds; final extension at 72°C for 10 minutes |
| Virulence genes (mrkD,rmpA,ybtS,entB,iutA,kfu, and allS) |
25: 10 PCR master mix, 2 DNA templates, 6 milliQ water, 0.5 of each primer |
95 °C for 15 minutes; 30 cycles of 94 °C for 30 seconds, 60 °C for 90 seconds, and 72 °C for 60 seconds; final extension at 72 °C for 10 minutes |
| Cephalosporin resistance (blaTEM,blaSHV, and blaCTX -M) |
10: 5 PCR master mix, 2 DNA templates, 2 milliQ water, 0.5 of each primer |
94 °C for 5 minutes; 37 cycles of 94 °C for 30 seconds, annealing for 30 seconds, 72 °C for 50 seconds; final extension at 72 °C for 5 minutes |
| Carbapenem resistance (blaNDM,blaKPC, and blaOXA) |
10: 5 PCR master mix, 2 DNA templates, 2 milliQ water, 0.5 of each primer |
94 °C for 4 minutes; 36 cycles of 94 °C for 30 seconds, annealing for 40 seconds, and 72 °C for 50 seconds; final extension at 72 °C for 5 minutes |
| Transport system genes (Ompk35,Ompk36,TolC,mdtk, and AcrAB) |
10: 5 PCR master mix, 2 DNA templates, 2 milliQ water, 0.5 of each primer |
95 °C for 15 minutes; 30 cycles of 94 °C for 30 seconds, specific annealing for 90 seconds, 72°C for 60 seconds; final extension at 72 °C for 10 minutes |
| Colistin resistance genes (mcr1-8) |
15: 5 PCR master mix, 2 DNA templates, 2 milliQ water, 0.5 of each primer |
94 °C for 15 minutes; 25 cycles of 94 °C for 30 seconds, annealing for 90 seconds, and 72 °C for 60 seconds; final extension at 72 °C for 10 minutes |
Note. PCR: Polymerase chain reaction.
Enterobacterial Repetitive Intergenic Consensus-Polymerase Chain Reaction
The diversity of K. pneumoniae isolates was assessed using ERIC-PCR with universal primers: forward (5′-ATGTAAGCTCCTGGGGATTCAC-3′) and reverse (5′-AAGTAAGTGACTGGGGTGAGCG-3′). The PCR mixture comprised 10 × PCR buffer, 10 pmol of each primer, 200 μM of deoxyribonucleotide triphosphate, 0.5 units of Taq polymerase, template DNA, and deionized water. Additionally, 35 cycles of denaturation at 94°C for 30 seconds, annealing at 47°C for 1 minute, and extension at 72 °C for 4 minutes were part of the PCR process, which began with an initial denaturation at 95 °C for 2.5 minutes. The final extension was conducted at 72 °C for 4 minutes. The resulting amplicons were separated on a 1.8% (wt/vol) agarose gel at 100 V for 35 minutes. Ultimately, the genetic relatedness of the strains was assessed by creating a dendrogram from the ERIC-PCR patterns using GelJ software, version 2.0.
Statistical Analysis
Descriptive statistics were determined using Microsoft Excel. Statistical significance was measured using chi-square analysis which was performed to assess associations between ERIC-PCR clusters, antibiotic resistance profiles, and virulence genes (P < 0.05).
Results
Antibiotic Susceptibility Testing
Phenotypic screening demonstrated that 78.74% (163/207) of the isolates were suspected ESBL producers, with 75.36% (156/207) confirmed positive. Moreover, most isolates were resistant to multiple antibiotics. The combined disc test identified 54.58% (113/207) as MBL producers (Figure 1a), with 61.94% (70/113) resistant to imipenem and 95.57% (108/113) resistant to meropenem. Overall, 95.65% (198/207) of the isolates were resistant to penicillin and ampicillin, while 77.77% (161/207) showed resistance to fluoroquinolones. In addition, sensitivity to chloramphenicol and fosfomycin was observed in 53.62% (111/207) and 54.0% (111/207) of the isolates, respectively (Figure 1b). Among the 207 uropathogenic Klebsiella isolates, 6.8% (14/207) were extensively drug-resistant (XDR), 68.1% (141/207) were MDR, 20.8% (43/207) were resistant to one or two classes, and 4.3% (9/207) were fully susceptible. Most XDR and MDR isolates were identified as K. pneumoniae subsp. pneumoniae (Figure 1c).
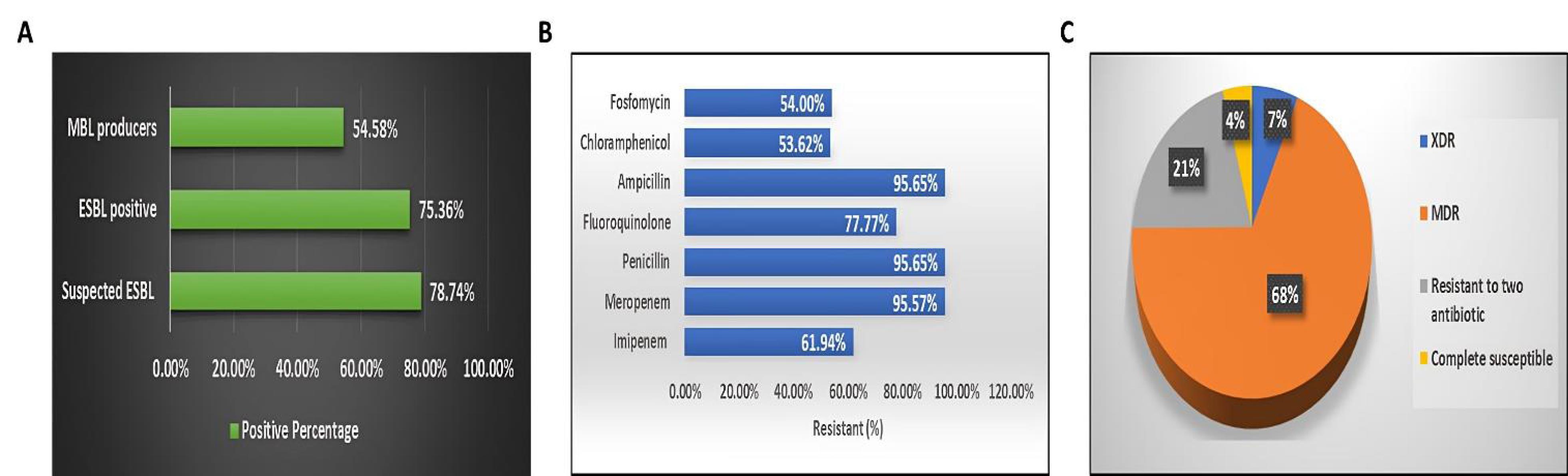
Figure 1.
(A) Identification of ESBL-Producing and MBL-Producing Klebsiella pneumoniae Isolates, (B) Percentage Distribution of Drug-Resistance Profiles Among the K. pneumoniae Isolates, and (C) Comparative Analysis of Multidrug-Resistant Isolates, Extensively Drug-Resistant Isolates, Isolates Resistant to Two Antibiotic Classes, and Susceptibility Rates of K. pneumoniae. Note. ESBL: Extended-spectrum beta-lactamase; MBL: Metallo-beta-lactamase
.
(A) Identification of ESBL-Producing and MBL-Producing Klebsiella pneumoniae Isolates, (B) Percentage Distribution of Drug-Resistance Profiles Among the K. pneumoniae Isolates, and (C) Comparative Analysis of Multidrug-Resistant Isolates, Extensively Drug-Resistant Isolates, Isolates Resistant to Two Antibiotic Classes, and Susceptibility Rates of K. pneumoniae. Note. ESBL: Extended-spectrum beta-lactamase; MBL: Metallo-beta-lactamase
Colistin Resistance
Initial screening using eosin methylene blue agar detected colistin resistance in 6.8% (14/207) of the isolates, all of which were K. pneumoniae subsp. pneumoniae. The minimum inhibitory concentration assay confirmed complete colistin resistance in all 14 isolates (CLSI-EUCAST). All 14 colistin-resistant isolates corresponded to the XDR category, consistent with their classification in the overall resistance analysis. None of the isolates belonged to the MDR-only group.
Capsular K1/K2 Serotype Detection
In a broader analysis of 207 isolates, molecular detection revealed that 4.3% (9/207) were positive for the K1 serotype, and 1.4% (3/207) were positive for the K2 serotype. Most K1 and K2 serotype isolates were K. pneumoniae subsp. pneumoniae (91.6%, 11/12). Among them, 66.7% (8/12) were MDR, and 83.3% (10/12) were strong biofilm producers (Figure 2).
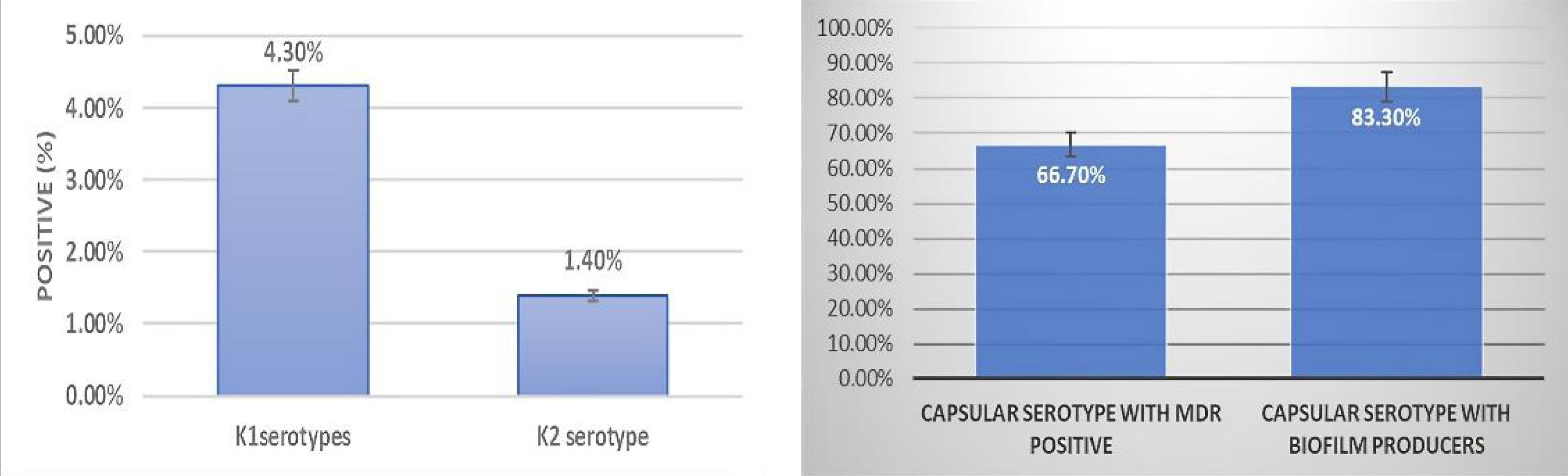
Figure 2.
(A) Graphs Representing the Proportion of Identified Capsule Serotypes (K1 and K2) Among the K. pneumoniae Isolates and (B) Classification of Capsule-Producing Klebsiella pneumoniae Strains Based on Their Multidrug-Resistance Status and Biofilm Production Capability
.
(A) Graphs Representing the Proportion of Identified Capsule Serotypes (K1 and K2) Among the K. pneumoniae Isolates and (B) Classification of Capsule-Producing Klebsiella pneumoniae Strains Based on Their Multidrug-Resistance Status and Biofilm Production Capability
Virulence Gene Detection
The detection of virulence genes in 207 isolates demonstrated varied profiles. The mrkD gene was present in 97.5% of isolates, indicating its widespread role as a fimbrial adhesin. In addition, the entB gene, involved in enterobactin synthesis, was found in 93.7% of isolates, highlighting its importance in iron acquisition. Further, the ybtS gene was detected in 42.9% of isolates, showing moderate prevalence. Furthermore, the kfu and iutA genes, both related to iron uptake, were observed in 21.7% of isolates each. Moreover, the allS gene, associated with allantoin metabolism, was present in 8.2% of isolates (Figure 3). The comparison of virulence gene distribution between MDR and non-MDR isolates, as well as between XDR and non-XDR isolates, revealed no statistically significant associations for any of the tested genes (all P > 0.05), emphasizing that the presence of virulence determinants is independent of MDR or XDR status.
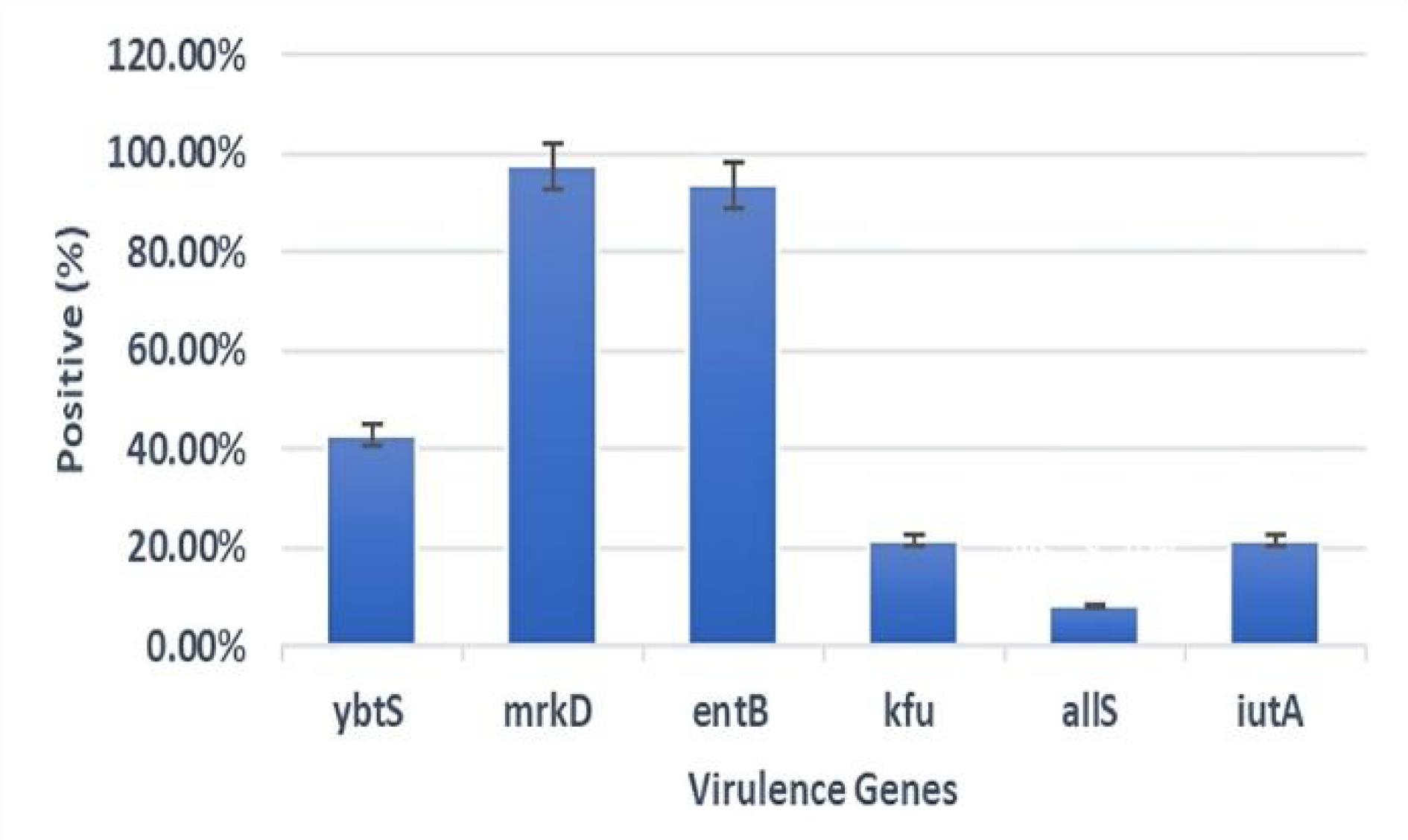
Figure 3.
Mapping and Prevalence of Virulence-Associated Genes Across the Clinical Isolates of K. pneumoniae. Note. This figure illustrates the identification of specific virulence genes and their frequency among the tested isolates
.
Mapping and Prevalence of Virulence-Associated Genes Across the Clinical Isolates of K. pneumoniae. Note. This figure illustrates the identification of specific virulence genes and their frequency among the tested isolates
Molecular Detection of Antibiotic Resistance Genes, Outer Membrane Porins, and Efflux Pumps
Genotypic analysis was performed on 24 isolates, comprising all XDR and colistin-resistant K. pneumoniae recovered in this study. Based on the results, 41.7% (10/24), 95.8% (23/24), and 70.8% (17/24) were positive for the blaTEM, blaSHV, and blaCTX -M genes, respectively. Additionally, 62.5% (15/24) and 33.3% (8/24) were positive for the blaNDM and blaOXA-48 genes, respectively, but none of them were positive for blaKPC. Additionally, none of the isolates carried any mcr gene variants. The molecular detection of outer membrane porins (omp) and efflux pump genes showed universal positivity for Ompk36. Eventually, the AcrAB, TolC, and Ompk35 genes were present in 95.8% of the isolates, and the mdtk gene was found in 79.2% (Figure 4).
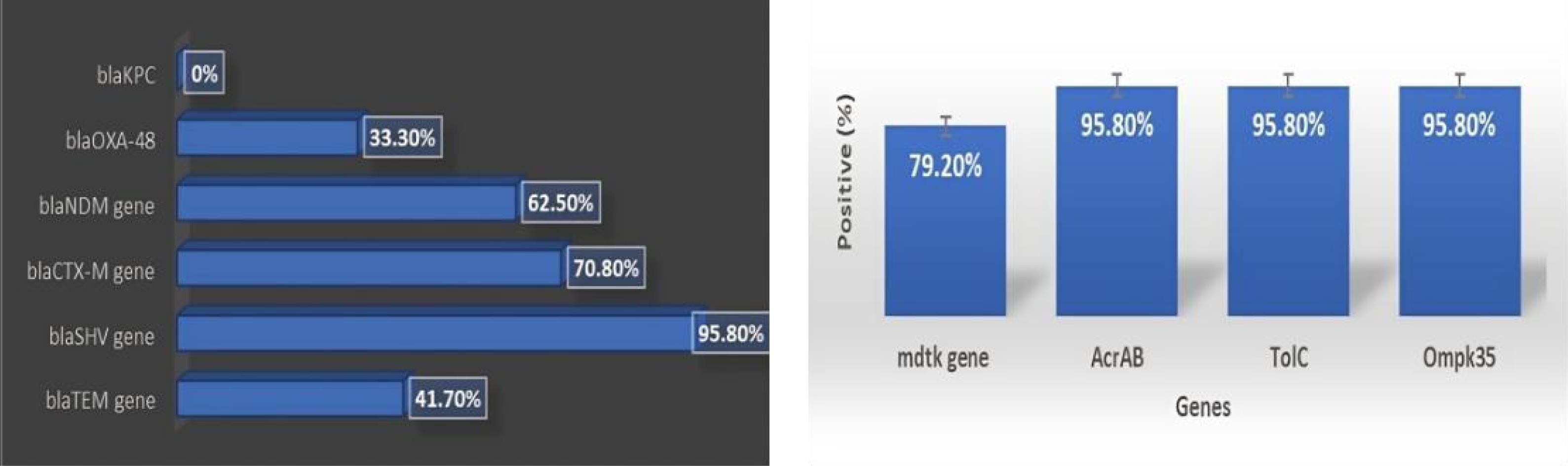
Figure 4.
(A) Molecular Profiling of Antibiotic Resistance Genes in Klebsiella pneumoniae Isolates and (B) Proportional Representation of Genes Associated With Outer Membrane Porins and Efflux Pumps in the Isolates
.
(A) Molecular Profiling of Antibiotic Resistance Genes in Klebsiella pneumoniae Isolates and (B) Proportional Representation of Genes Associated With Outer Membrane Porins and Efflux Pumps in the Isolates
Enterobacterial Repetitive Intergenic Consensus-Polymerase Chain Reaction Analysis
A total of 207 DNA samples were collected for analysis. Of them, 24 samples (12%) had no ERIC pattern, but 183 samples (88%) had ERIC patterns. The online program GelJ (version 2.0) was used to perform the ERIC-PCR analysis. One to eight bands, ranging in size from 50 bp to 1500 bp, were found in each sample by the ERIC-PCR gel analysis (Figure 5). Although 88% of the isolates showed ERIC patterns, clustering analysis revealed no significant association between ERIC profiles and resistance categories. To ascertain the genetic relatedness of the strains, a dendrogram and clustering analysis were created using the ERIC patterns (Figure 6).
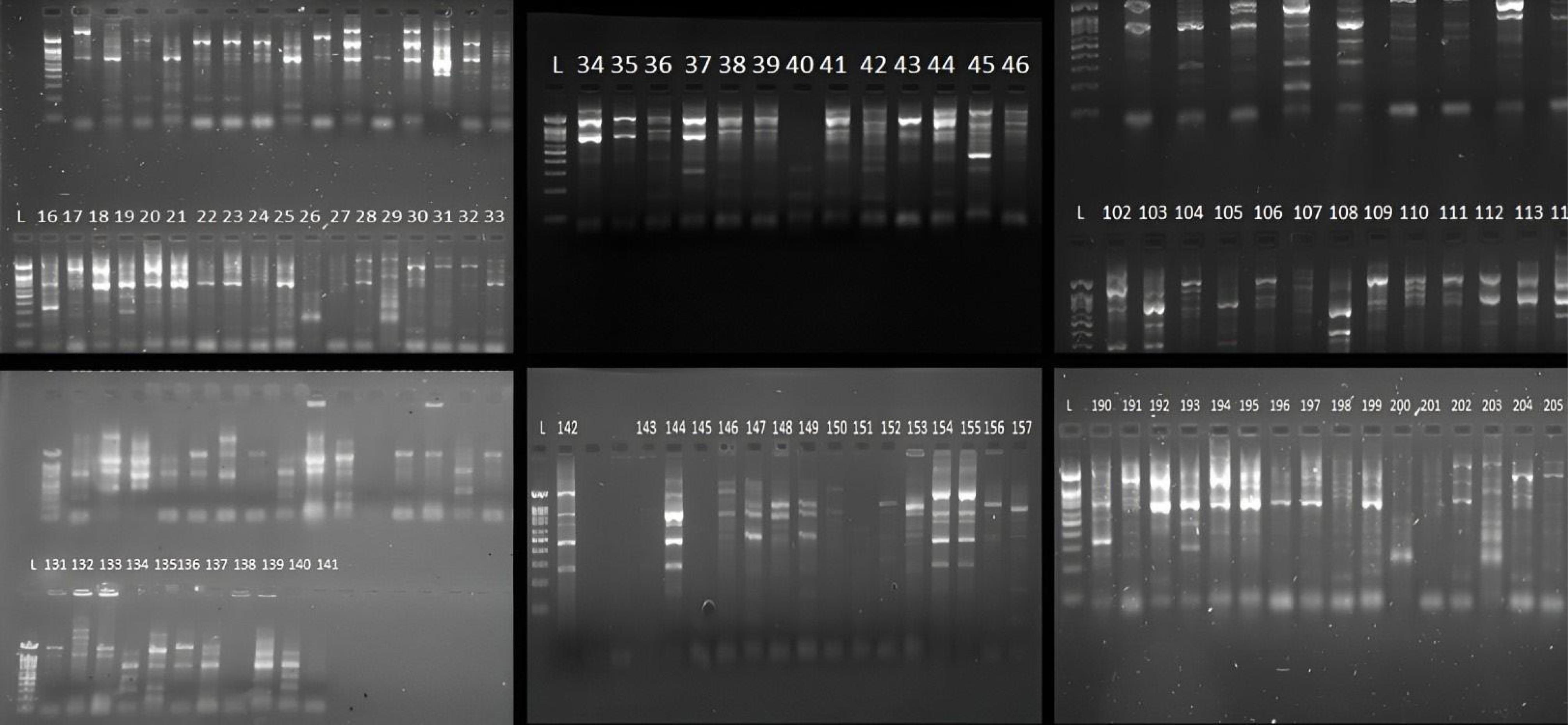
Figure 5.
Gel Electrophoresis Results of Enterobacterial Repetitive Intergenic Consensus-Polymerase Chain Reaction Fingerprinting, Showcasing Genetic Diversity Among Klebsiella pneumoniae Isolates
.
Gel Electrophoresis Results of Enterobacterial Repetitive Intergenic Consensus-Polymerase Chain Reaction Fingerprinting, Showcasing Genetic Diversity Among Klebsiella pneumoniae Isolates
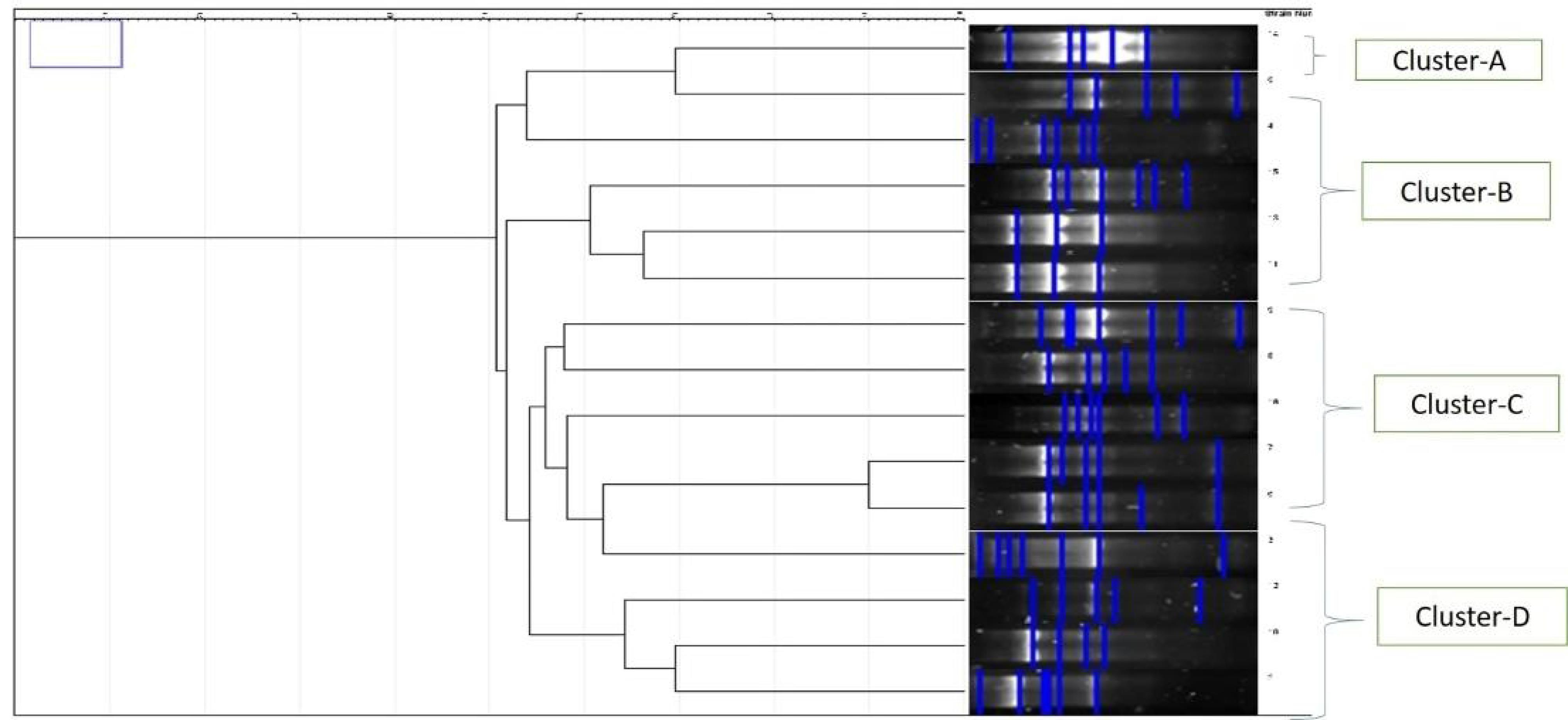
Figure 6.
Dendrogram Illustrating the Clustering of Klebsiella pneumoniae Isolates Based on Enterobacterial Repetitive Intergenic Consensus-Polymerase Chain Reaction Fingerprinting, Highlighting Genetic Relationships and Diversity Among the Isolates
.
Dendrogram Illustrating the Clustering of Klebsiella pneumoniae Isolates Based on Enterobacterial Repetitive Intergenic Consensus-Polymerase Chain Reaction Fingerprinting, Highlighting Genetic Relationships and Diversity Among the Isolates
Discussion
With an emphasis on XDR strains, the molecular mechanisms behind antibiotic resistance in MDR K. pneumoniae strains, as well as their potential for pathogenicity, were examined in this study. Due to the extensive use of antimicrobial drugs, the incidence of MDR K. pneumoniae infections has increased throughout the past ten years (18). Considering that there are few effective antibiotic treatment options available, this trend presents a serious health risk. According to the findings, K. pneumoniae (98.1%) was the most common species among the uropathogenic Klebsiella isolates, which is in line with the results of previous research, indicating that K. pneumoniae was the primary perpetrator of UTIs (19,20). In this investigation, 76.8% of the Klebsiella isolates produced ESBLs, which is extremely high in comparison to previous investigations. Our findings contradict those of a prior South Indian study, demonstrating that 35.7% of the Klebsiella isolates were resistant to carbapenem (21). Colistin resistance was observed in 6.8% of K. pneumoniae isolates in our study, which does not match the results of other studies, reporting colistin resistance in 2.1%, 11.1%, 18.5%, and 19% of the K. pneumoniae isolates. Nearly 25% of XDR and colistin-resistant K. pneumoniae isolates were found to be significant ESBL producers using the E-strip MIC test, which is consistent with previous findings. MBL, AmpC, or KPC were not produced by any of the XDR isolates in our study, which conforms to the findings of other studies (22,23).
In this study, the K1-4.3% serotype was shown to be more common than the K2-1.4% serotype, which corroborates the results of a study conducted by Yeh et al, in which the K1 serotype (46.6%) was more common than the K2 serotype (20.5%) (24). Our findings confirmed high prevalence rates of virulence genes mrkD (98%) and entB (94%) in K. pneumoniae isolates, aligning with the findings of Rastegar et al (25), demonstrating 94.5% and 95%, respectively, for K1 and K2 serotypes. These findings are also in conformity with those of Compain et al (10). None of our isolates tested positive for the rmpA gene, and most did not exhibit the hypermucoviscous phenotype. This supports the findings, which associated the rmpA gene with hypermucoviscosity in K. pneumoniae. In a study conducted by Mirzaie and Ranjbar, the virulence-associated gene entB showed 75% resistance to ciprofloxacin in 80% of the isolates, and the efflux pump genes (AcrAB, TolC, and mdtk) were observed in 41%, 33%, and 26% of the strains (26), which matches our findings. Despite the high prevalence of mrkD and entB and the moderate/low frequencies of other virulence genes, our statistical analysis revealed no significant association between virulence gene distribution and MDR or XDR status (all P > 0.05). This indicates that virulence determinants are broadly distributed across resistant and susceptible isolates. The majority of ESBL-producing isolates were positive for the blaSHV gene (95.8%), which is in line with the results of other studies that identified the blaSHV gene in 87% and 62% of the isolates, respectively. Similarly, our results demonstrated that 71% of the isolates had the blaCTX -M gene, which is consistent with the findings of other studies, indicating that 44.6%, 69%, and 89.4% of the isolates carried the blaCTX -M gene (27,28). ESBL genes blaTEM, blaSHV, and blaCTX -M (29.79%, 27.66%, and 17.02%) in ESBL-producing isolates were found positive, along with K1 and K2 serotypes and the kfu virulence gene among the isolates in another study by Zhong et al (28). Based on our findings, 41.7% of the isolates had the blaTEM gene, while a previous study reported that 36% of the isolates had the blaTEM gene (28). The presence of the blaCTX -M, blaTEM, blaOXA, and blaSHV (100%, 97%, 86%, and 83%) ESBL genes in similar genetic contexts has been confirmed in a South African study. The blaNDM gene was found in 62.5% of the carbapenemase-producing K. pneumoniae isolates compared to 7.1% and 37% in previous studies. In a study in Bangladesh, the analysis showed that the samples carried 64% and 38% blaNDM and blaSHV genes, and 19% carried both genes (28). The blaOXA-48 gene was identified in 33% of the carbapenemase-producing K. pneumoniae isolates. Nonetheless, reports from other studies showed that 6%, 9%, 67%, and 71% of the isolates carried blaOXA-48, respectively. In our study, Ompk36 was detected in all the isolates (100%), followed by Ompk35 (96%), which conforms to the results of a study performed in Iran (28). The frequency of the AcrAB gene (96%) was higher than that of the mdtk gene (73%) in our XDR and colistin-resistant K. pneumoniae isolates, which is consistent with the results of a study conducted in Egypt (28). In the present study, 96% of XDR and colistin-resistant K. pneumoniae isolates were positive for the TolC gene, which corroborates the findings of the Brazilian study, where, in addition to the TolC gene, efflux pump-related genes AcrAB (100%), mdtk (88%), Ompk35 (60%), and Ompk36 (28%) were also found positive. Among ESBL producers, blaKPC (100%), blaTEM (100%), blaSHV (96%), blaOXA (84%), and blaCTX -M (72%) amr genes were the carriers. The genotyping results of colistin-resistant K. pneumoniae isolates revealed that the absence of a plasmid encodes resistance genes (mcr). In the absence of mcr genes, the presence of the chromosomally mediated gene mgrB may be linked to colistin resistance in K. pneumoniae. According to earlier research, mutations in the mgrB gene were the most frequent sources of colistin resistance in K. pneumoniae. In this study, 207 DNA samples were analyzed using ERIC-PCR to assess genetic diversity and relatedness among bacterial strains. Our results showed that 183 samples (88%) exhibited ERIC patterns, while 24 samples (12%) did not display these patterns, which aligns with the results of Meacham et al (29), demonstrating variations in the genetic diversity of bacterial isolates by ERIC-PCR. They successfully used ERIC-PCR to construct a phylogenetic tree for Klebsiella, highlighting the method’s effectiveness. Similarly, Barus et al identified 16 clusters from 61 Klebsiella spp. isolates based on ERIC-PCR profiles, revealing significant genetic diversity within K. pneumoniae (30), which is consistent with our findings, underscoring the utility of ERIC-PCR in discerning genetic diversity. The 1 to 8 bands per sample observed in our analysis are comparable to the diversity found in studies performed by Meacham et al (29) and Barus et al (30), further validating the robustness of ERIC-PCR for genetic analysis. Moreover, the dendrogram and clustering analysis in our study revealed distinct clusters, indicating significant genetic diversity among the bacterial strains, which corroborates the clustering patterns observed in the study by Barus et al (30), where different ERIC profiles described the genetic diversity within Klebsiella spp. In conclusion, our study, along with those of Meacham et al (29) and Barus et al (30), demonstrated the effectiveness of ERIC-PCR in revealing genetic diversity and relatedness among bacterial isolates. In accordance with our statistical analysis, no significant associations were found between ERIC-PCR clustering, antibiotic resistance, and virulence gene distribution (P > 0.05). This suggests that resistance and virulence determinants were not confined to specific clonal groups but rather dispersed across genetically diverse isolates. The lack of correlation may reflect the multifactorial nature of resistance and virulence acquisition in K. pneumoniae, where mobile genetic elements and horizontal gene transfer play a key role. One limitation of this study was that multivariate approaches (e.g., logistic regression) could not be applied due to the uneven distribution of virulence genes, some being nearly universal (mrkD and entB) and others rare (rmpA and allS). This imbalance restricted the robustness of regression modeling and may have limited deeper exploration of the predictors of MDR/XDR phenotypes. Future large-scale studies integrating whole genome sequencing and advanced statistical approaches may better delineate these complex relationships. Better resolution of such correlations may be possible with larger sample sizes and whole-genome-based methodologies. These consistent findings reinforce the method’s reliability and robustness, making it a valuable tool for genetic analysis in microbial populations.
Conclusion
Our findings revealed the complex genetic landscape of K. pneumoniae, with multiple resistance and virulence factors coexisting, complicating infection control and treatment. The diverse virulence profiles highlighted the pathogen’s ability to employ various pathogenic strategies. It is essential to comprehend these elements in order to create prevention and treatment plans that are effective. Moreover, continuous surveillance and molecular characterization are essential for informed clinical management and targeted therapies. Addressing the burden of MDR and XDR K. pneumoniae requires stringent infection control and the development of novel therapeutic options. Accordingly, future research should focus on the genetic diversity and evolution of resistance and virulence determinants.
Acknowledgments
The authors of this manuscript thank the Department of Health Research-Multidisciplinary Research Units at Dr. ALM PG Institute of Basic Medical Sciences, the University of Madras, for valuable assistance in this research.
Competing Interests
The authors declare that they have no conflict of interests.
Funding
This research received no specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Supplementary Files
The supplementary file contains Table S1.
(pdf)
References
- Mody L, Juthani-Mehta M. Urinary tract infections in older women: a clinical review. JAMA 2014; 311(8):844-54. doi: 10.1001/jama.2014.303 [Crossref] [ Google Scholar]
- Farajnia S, Alikhani MY, Ghotaslou R, Naghili B, Nakhlband A. Causative agents and antimicrobial susceptibilities of urinary tract infections in the northwest of Iran. Int J Infect Dis 2009; 13(2):140-4. doi: 10.1016/j.ijid.2008.04.014 [Crossref] [ Google Scholar]
- Podschun R, Ullmann U. Klebsiella spp as nosocomial pathogens: epidemiology, taxonomy, typing methods, and pathogenicity factors. Clin Microbiol Rev 1998; 11(4):589-603. doi: 10.1128/cmr.11.4.589 [Crossref] [ Google Scholar]
- Lopes AC, Rodrigues JF, Clementino MB, Miranda CA, Nascimento AP, de Morais Júnior MA. Application of PCR ribotyping and tDNA-PCR for Klebsiella pneumoniae identification. Mem Inst Oswaldo Cruz 2007; 102(7):827-32. doi: 10.1590/s0074-02762007005000113 [Crossref] [ Google Scholar]
- Ballén V, Gabasa Y, Ratia C, Ortega R, Tejero M, Soto S. Antibiotic resistance and virulence profiles of Klebsiella pneumoniae strains isolated from different clinical sources. Front Cell Infect Microbiol 2021; 11:738223. doi: 10.3389/fcimb.2021.738223 [Crossref] [ Google Scholar]
- Riwu KH, Effendi MH, Rantam FA, Khairullah AR, Widodo A. A review: virulence factors of Klebsiella pneumonia as emerging infection on the food chain. Vet World 2022; 15(9):2172-9. doi: 10.14202/vetworld.2022.2172-2179 [Crossref] [ Google Scholar]
- Paczosa MK, Mecsas J. Klebsiella pneumoniae: going on the offense with a strong defense. Microbiol Mol Biol Rev 2016; 80(3):629-61. doi: 10.1128/mmbr.00078-15 [Crossref] [ Google Scholar]
- Cubero M, Marti S, Domínguez M, González-Díaz A, Berbel D, Ardanuy C. Hypervirulent Klebsiella pneumoniae serotype K1 clinical isolates form robust biofilms at the air-liquid interface. PLoS One 2019; 14(9):e0222628. doi: 10.1371/journal.pone.0222628 [Crossref] [ Google Scholar]
- Holt KE, Wertheim H, Zadoks RN, Baker S, Whitehouse CA, Dance D. Genomic analysis of diversity, population structure, virulence, and antimicrobial resistance in Klebsiella pneumoniae, an urgent threat to public health. Proc Natl Acad Sci U S A 2015; 112(27):E3574-81. doi: 10.1073/pnas.1501049112 [Crossref] [ Google Scholar]
- Compain F, Babosan A, Brisse S, Genel N, Audo J, Ailloud F. Multiplex PCR for detection of seven virulence factors and K1/K2 capsular serotypes of Klebsiella pneumoniae. J Clin Microbiol 2014; 52(12):4377-80. doi: 10.1128/jcm.02316-14 [Crossref] [ Google Scholar]
- Nakamura-Silva R, Oliveira-Silva M, Furlan JPR, Stehling EG, Miranda CE, Pitondo-Silva A. Characterization of multidrug-resistant and virulent Klebsiella pneumoniae strains belonging to the high-risk clonal group 258 (CG258) isolated from inpatients in northeastern Brazil. Arch Microbiol 2021; 203(7):4351-9. doi: 10.1007/s00203-021-02425-0 [Crossref] [ Google Scholar]
- Paterson DL, Bonomo RA. Extended-spectrum beta-lactamases: a clinical update. Clin Microbiol Rev 2005; 18(4):657-86. doi: 10.1128/cmr.18.4.657-686.2005 [Crossref] [ Google Scholar]
- Shakib P, Taheri Kalani M, Ramazanzadeh R, Ahmadi A, Rouhi S. Molecular detection of virulence genes in Klebsiella pneumoniae clinical isolates from Kurdistan province, Iran. Biomed Res Ther 2018; 5(8):2581-9. doi: 10.15419/bmrat.v5i8.467 [Crossref] [ Google Scholar]
- Kundu J, Kansal S, Rathore S, Kaundal M, Angrup A, Biswal M. Evaluation of ERIC-PCR and MALDI-TOF as typing tools for multidrug resistant Klebsiella pneumoniae clinical isolates from a tertiary care center in India. PLoS One 2022; 17(11):e0271652. doi: 10.1371/journal.pone.0271652 [Crossref] [ Google Scholar]
- Heamchandsaravanan AR, Shanmugam K, Perumal D, Shankar D, Kalpana S, Dhandapani P. Loop-mediated isothermal amplification (LAMP) assay targeting STY2879 gene for rapid detection of Salmonella enterica serovar Typhi in blood. Res J Pharm Technol 2024; 17(5):2087-92. doi: 10.52711/0974-360x.2024.00330 [Crossref] [ Google Scholar]
- Mitra S, Mukherjee S, Naha S, Chattopadhyay P, Dutta S, Basu S. Evaluation of co-transfer of plasmid-mediated fluoroquinolone resistance genes and bla(NDM) gene in Enterobacteriaceae causing neonatal septicaemia. Antimicrob Resist Infect Control 2019; 8:46. doi: 10.1186/s13756-019-0477-7 [Crossref] [ Google Scholar]
- Wasfi R, Elkhatib WF, Ashour HM. Molecular typing and virulence analysis of multidrug resistant Klebsiella pneumoniae clinical isolates recovered from Egyptian hospitals. Sci Rep 2016; 6:38929. doi: 10.1038/srep38929 [Crossref] [ Google Scholar]
- Hou XH, Song XY, Ma XB, Zhang SY, Zhang JQ. Molecular characterization of multidrug-resistant Klebsiella pneumoniae isolates. Braz J Microbiol 2015; 46(3):759-68. doi: 10.1590/s1517-838246320140138 [Crossref] [ Google Scholar]
- Shakya P, Shrestha D, Maharjan E, Sharma VK, Paudyal R. ESBL production among E coli and Klebsiella spp causing urinary tract infection: a hospital-based study. Open Microbiol J 2017; 11:23-30. doi: 10.2174/1874285801711010023 [Crossref] [ Google Scholar]
- Devrim F, Serdaroğlu E, Çağlar İ, Oruç Y, Demiray N, Bayram N. The emerging resistance in nosocomial urinary tract infections: from the pediatrics perspective. Mediterr J Hematol Infect Dis 2018; 10(1):e2018055. doi: 10.4084/mjhid.2018.055 [Crossref] [ Google Scholar]
- Dillirani V, Suresh R. A study on prevalence and antimicrobial resistance pattern of urinary Klebsiella pneumoniae in a tertiary care centre in South India. Int J Curr Microbiol Appl Sci 2018; 7(4):969-73. doi: 10.20546/ijcmas.2018.704.103 [Crossref] [ Google Scholar]
- Bartolini A, Frasson I, Cavallaro A, Richter SN, Palù G. Comparison of phenotypic methods for the detection of carbapenem non-susceptible Enterobacteriaceae. Gut Pathog 2014; 6:13. doi: 10.1186/1757-4749-6-13 [Crossref] [ Google Scholar]
- Santino I, Bono S, Nuccitelli A, Martinelli D, Petrucci C, Alari A. Microbiological and molecular characterization of extreme drug-resistant carbapenemase-producing Klebsiella pneumoniae isolates. Int J Immunopathol Pharmacol 2013; 26(3):785-90. doi: 10.1177/039463201302600325 [Crossref] [ Google Scholar]
- Yeh KM, Kurup A, Siu LK, Koh YL, Fung CP, Lin JC. Capsular serotype K1 or K2, rather than magA and rmpA, is a major virulence determinant for Klebsiella pneumoniae liver abscess in Singapore and Taiwan. J Clin Microbiol 2007; 45(2):466-71. doi: 10.1128/jcm.01150-06 [Crossref] [ Google Scholar]
- Rastegar S, Moradi M, Kalantar-Neyestanaki D, Golabi Dehdasht A, Hosseini-Nave H. Virulence factors, capsular serotypes and antimicrobial resistance of hypervirulent Klebsiella pneumoniae and classical Klebsiella pneumoniae in southeast Iran. Infect Chemother. 2019. doi: 10.3947/ic.2019.0027.
- Mirzaie A, Ranjbar R. Antibiotic resistance, virulence-associated genes analysis and molecular typing of Klebsiella pneumoniae strains recovered from clinical samples. AMB Express 2021; 11(1):122. doi: 10.1186/s13568-021-01282-w [Crossref] [ Google Scholar]
- Nirwati H, Sinanjung K, Fahrunissa F, Wijaya F, Napitupulu S, Hati VP. Biofilm formation and antibiotic resistance of Klebsiella pneumoniae isolated from clinical samples in a tertiary care hospital, Klaten, Indonesia. BMC Proc 2019; 13(Suppl 11):20. doi: 10.1186/s12919-019-0176-7 [Crossref] [ Google Scholar]
- Zhong XS, Li YZ, Ge J, Xiao G, Mo Y, Wen YQ. Comparisons of microbiological characteristics and antibiotic resistance of Klebsiella pneumoniae isolates from urban rodents, shrews, and healthy people. BMC Microbiol 2020; 20(1):12. doi: 10.1186/s12866-020-1702-5 [Crossref] [ Google Scholar]
- Meacham KJ, Zhang L, Foxman B, Bauer RJ, Marrs CF. Evaluation of genotyping large numbers of Escherichia coli isolates by enterobacterial repetitive intergenic consensus-PCR. J Clin Microbiol 2003; 41(11):5224-6. doi: 10.1128/jcm.41.11.5224-5226.2003 [Crossref] [ Google Scholar]
- Barus T, Hanjaya I, Sadeli J, Lay BW, Suwanto A, Yulandi A. Genetic diversity of Klebsiella spp isolated from tempe based on enterobacterial repetitive intergenic consensus-polymerase chain reaction (ERIC-PCR). Hayati 2013; 20(4):171-6. doi: 10.4308/hjb.20.4.171 [Crossref] [ Google Scholar]