Avicenna Journal of Clinical Microbiology and Infection. 12(4):195-203.
doi: 10.34172/ajcmi.3678
Original Article
Impact of Multiplicity of Infection (MOI) on In-Vitro Phage Therapy Against Xanthomonas axonopodis pv. citri
Mohammad Irfan Khan Conceptualization, Formal analysis, Investigation, Writing – original draft, 1 
Shahzeb Javed Conceptualization, Formal analysis, Investigation, Writing – original draft, 1
Mujaddad Ur Rehman Conceptualization, Formal analysis, Funding acquisition, Investigation, Writing – original draft, 1, *
Muhammad Rizwan Conceptualization, Formal analysis, Investigation, Writing – original draft, 2
Ibrar Khan Data curation, Validation, Writing – review & editing, 1
Azam Hayat Data curation, Validation, Writing – review & editing, 1
Iqbal Ahmad Alvi Data curation, Validation, Writing – review & editing, 3
Amir Sasan Mozaffari Nejad Conceptualization, Data curation, Formal analysis, Investigation, Validation, Writing – original draft, Writing – review & editing, 4, 5, *
Author information:
1Department of Microbiology, Abbottabad University of Science & Technology, 22010 Havelian, Pakistan
2USPCAS-W Mehran University of Engineering and Technology Jamshoro, Sindh, Pakistan
3Department of Microbiology, Hazara University, 21120 Mansehra, Pakistan
4Bio Environmental Health Hazards Research Center, Jiroft University of Medical Sciences, Jiroft, Iran
5Infectious Disease and Tropical Medicine Research Center, Isfahan University of Medical Sciences, Isfahan, Iran
Abstract
Background: Antimicrobial resistance (AMR) is an escalating global health concern, with the rapid emergence of multidrug-resistant (MDR) pathogens outstripping the development of new antibiotics. Bacteriophages (phages), which specifically target and lyse bacteria, represent a promising alternative. This study investigated the therapeutic potential of phages against Xanthomonas axonopodis pv. citri, a significant bacterial pathogen causing citrus canker and considerable economic losses worldwide.
Methods: Phages were isolated from diverse environmental sources (e.g., soil and citrus fruit samples) and screened for lytic activity against X. axonopodis pv. citri. Phage purification and titer determination were performed using the double-layer agar method. The ability of phages to suppress bacterial growth was assessed in vitro via optical density (OD₆₀₀) and colony-forming unit (CFU) counts over 24 hours across a range of multiplicities of infection (MOIs).
Results: The isolated phages exhibited a narrow host range and high stability at 7°C, with an average titer of 201 plaques per plate upon purification. In vitro assays confirmed that the phages successfully suppressed the growth of X. axonopodis pv. citri, demonstrating strong lytic activity and significant reduction in bacterial loads. Phages 1P, 2P, 3P, and 5P showed varying degrees of effectiveness, with notable suppression at MOIs of 0.01, 10, and 100. Phage 3P provided sustained bacterial suppression at MOI 10, while phage 2P showed consistent activity across all tested MOIs. Despite observations of transient bacterial resistance, all phages maintained their antibacterial efficacy.
Conclusion: Our findings underscore the therapeutic potential of bacteriophages as an effective and sustainable alternative to antibiotics for controlling X. axonopodis pv. citri. The demonstrated lytic activity and bacterial suppression, even at varying MOIs, suggest that phage therapy offers a viable strategy against antibiotic-resistant strains. Furthermore, the cost-effectiveness of phage therapy provides a compelling rationale for its expanded consideration in addressing the global antibiotic resistance crisis within the broader context of healthcare economics.
Keywords: Antibiotics, Antimicrobial resistance, Bacteriophage, Phage therapy, MOI
Copyright and License Information
© 2025 The Author(s); Published by Hamadan University of Medical Sciences.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (
https://creativecommons.org/licenses/by/4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Please cite this article as follows: Khan MI, Javed S, Rehman MU, Rizwan M, Khan I, Hayat A, et al. Impact of multiplicity of infection (MOI) on In-vitro phage therapy against Xanthomonas axonopodis pv. citri. Avicenna J Clin Microbiol Infect. 2025;12(4):195-203. doi:10.34172/ajcmi.3678
Introduction
Antibiotics are a class of medications administered in controlled doses to eliminate or inhibit the growth of microorganisms, particularly bacteria, and resistance to antibiotics is the ability of bacterial cells to withstand, survive, and grow in the presence of antibiotics, resulting in the survival of bacteria rather than destruction (1-4). Antibiotic resistance has emerged as a significant global challenge, leading to infections by multidrug-resistant (MDR) pathogens that are increasingly difficult to treat. Bacterial cells acquire resistance through various mechanisms, including genetic alterations, such as DNA mutations, and the acquisition of drug-resistance genes, often exacerbated by the improper use of antibiotics. Consequently, bacteria employ diverse strategies to counteract the effects of these therapeutic agents (5-8). It is a fact that the number of MDR bacteria has been rising as a result of the widespread and excessive use of antibiotics. Misuse and overuse of antibiotics have also increased the emergence of MDR bacteria (6,9). Annually, a considerable number of individuals are admitted to hospitals worldwide, and this figure continues to rise. Patients in intensive care units (ICUs) face a particularly high risk of infections. These infections are frequently caused by MDR microorganisms, posing a significant threat to patient outcomes and public health (10,11).
The use of antibiotics in agriculture and animal husbandry, alongside human medicine, is linked to the rising concern over antimicrobial resistance (AMR). High levels of antibiotic consumption in animal husbandry are particularly notable in countries like China, the United States, and Brazil (12-14). Citrus canker, caused by Xanthomonas axonopodis pv. citri, poses a significant threat to the citrus industry. The disease, which has become more severe in the last decade due to resistant Xanthomonas strains, leads to major crop losses worldwide (15). Therefore, there is an urgent need to solve this global problem.
Bacteriophages (phages) are viruses that specifically infect and lyse bacteria. Their high host-specificity and lack of threat to eukaryotic cells make them a promising therapeutic alternative (16,17). The therapeutic application of bacteriophages was first proposed by Twort and d’Herelle in the early 20th century (18). Evidence suggests that phages and their encoded proteins can be effectively utilized to treat bacterial infections, especially in scenarios where conventional antibiotic therapy is ineffective (19). The phages are highly host-specific. They replicate inside the host bacterium and multiply in large numbers to maintain their population for critical times (20). For almost a century, phages have been used in therapeutic settings to treat bacterial diseases (21,22). They appeared to be effective in the treatment of chronic infections (23,24), wound infections, or lung-associated infections in various studies (25,26). According to the WHO report on bacteriophage therapy, only in the Eliava Phage Therapy Center in Tbilisi, Georgia, 5256 patients were treated using bacteriophage therapy across various medical specialties between 2019 and 2023 (27-30). Phage therapy is a century-old treatment targeting bacterial infection, which was used to treat over 6300 patients in various clinical specialties from 1922 to 2022 and was implemented across the world in Europe, America, Asia, and Africa (31).
Bacteriophage therapy, despite its potential, faces challenges that limit its efficacy, such as the need for an optimal multiplicity of infection (MOI) to avoid poor eradication or undesirable immune responses. However, phages offer key advantages, including their ability to replicate at the infection site and their high host specificity. Therefore, this study aimed to isolate and characterize lytic bacteriophages specific to Xanthomonas axonopodis pv. citri and to identify appropriate therapeutic doses. This was achieved by determining the optimal MOI for these phages, aiming for effective bacterial clearance while maintaining phage stability and minimizing potential drawbacks.
Materials and Methods
Collection of Bacterial Strain
An environmental isolate of X. axonopodis pv. citri (accession number: MH106434) was provided by Dr. Mujaddad-ur-Rehman (Department of Microbiology, Abbottabad University of Science and Technology, Pakistan). The bacterial strain was utilized as a host for bacteriophage isolation and characterization.
Bacteriophage Isolation and Propagation
To isolate bacteriophages, soil samples were collected from Khanpur region in Haripur. The collection and processing were performed using a previously reported method with slight modification (32,33). Through this refined procedure, a bacteriophage was successfully isolated from these soil samples. For this purpose, 10 g soil samples were collected from citrus orchards exhibiting citrus canker symptoms. These samples were transferred to conical flasks and left undisturbed for 24 hours to facilitate the sedimentation of heavy particles. Then, the soil was suspended in 90 mL of SM buffer [100 mM NaCl, 8 mM MgSO₄, 50 mM Tris-HCl (pH 7.5)] and thoroughly vortexed for 10 minutes to release phages into the solution. After 24 hours, the clear supernatant was filtered using a 0.22 µm sterile syringe filter and collected in a separate sterile Falcon tube. For phage enrichment, 10 mL of the filtered supernatant was combined with 10 mL of an overnight culture of X. axonopodis pv. citri. The resulting enriched filtrate was subsequently assessed using the spot assay.
Assessment of Bacteriophage by Spot Assay
Bacteriophage presence was confirmed using the spot assay, as previously described (34,35). Phages showing robust lytic activity on the most susceptible host strains were then selected. These active filtered phage isolates were subsequently advanced for further analysis.
Purification of Bacteriophages
Initially, serial dilutions of the phage suspension were prepared for purification. Each dilution received an addition of 100 µL of an overnight bacterial culture (X. axonopodis pv. citri). The resulting mixtures were subsequently incubated at 37 °C for 5 minutes. After pouring the mixture onto an LB agar plate, 3-4 mL of semi-solid LB medium was added to form an overlay. Each Petri plate was then gently rotated to ensure thorough and even distribution of the overlay. Afterwards, the plates were incubated at 37 °C for 24 hours. After overnight incubation, small clear plaques, indicative of phage formation, were observed on the LB agar plates. Using a sterile micropipette tip, an isolated plaque was picked from the LB agar plate. This plaque-picking process was repeated two to three times to ensure the purification of the phage isolate. The purified phages were then stored in SM buffer supplemented with 0.002% (w/v) gelatin for short-term storage at 4 °C. For long-term preservation, phages were stored at −80 °C with the addition of 7% (v/v) dimethyl sulfoxide (DMSO) (32).
Phage Titer Determination
Titration of phages was done using the double-layer agar method described by Adams (36). The phage stock solution was serially diluted. Briefly, 100 μL of each dilution was transferred into different sterile test tubes. Then, 100 µL of bacterial culture was added to each test tube and kept at room temperature for 10 minutes. Afterwards, 3 mL of 0.7% top agar was added. The mixture was carefully poured on fresh nutrient agar plates and kept for 10-15 minutes to allow the top agar layer to solidify. Then, the mixture was incubated at 37 ºC for 24 hours, and the plates were checked for plaque formation. Finally, the plates with clear plaques were selected for phage titer determination.
(PFU/mL) = number of plaques / D × V
D = dilution factor
V = volume of diluted virus
Determination of Bacterial Growth Curve
Briefly, 50 mL of LB broth was aliquoted into two 250 mL Erlenmeyer flasks. One flask was inoculated with a fresh culture of X. axonopodis pv. citri, while the other served as an uninoculated control. Both flasks were incubated at 37 °C. Optical density measurements at 600 nm (OD₆₀₀) were performed every 30 minutes using a spectrophotometer, commencing after two hours of incubation. After 24 hours of incubation, samples were serially diluted and plated on agar to determine colony-forming units (CFUs). Both CFU counts and OD₆₀₀ measurements were subsequently plotted over time to characterize bacterial growth (37).
Determination of Bacteriophage MOI for Bacterial Growth Reduction
To assess bacterial growth and evaluate the effectiveness of phage infection, a bacterial reduction assay was performed, as described previously (38). This assay quantifies bacterial growth by measuring bacterial viability over time. For this assay, a standardized overnight culture of X. axonopodis pv. citri at a concentration of 3 × 107 CFU/mL was utilized. Four selected bacteriophage isolates (1P, 2P, 3P, and 5P) were individually tested. Experiments were conducted in a 96-well plate format with varying MOIs, defined as the initial ratio of phages to bacterial cells.
The plate setup was as follows:
Column 1: Contained 100 µL of LB broth and served as a sterile negative control.
Column 2: Inoculated with 3 × 107 CFU/mL of bacterial culture alone, serving as a positive control for bacterial growth.
Columns 3 to 8: Contained bacterial culture mixed with phages at MOI values of 0.001, 0.01, 0.1, 1, 10, and 100, respectively, to investigate the effect of varying phage concentrations on bacterial growth.
Upon completion of the plate setup, the plates were incubated at 37 °C. At 2, 4, 8, 12, 16, 20, and 24 hours post-incubation, 10 µL samples were collected from each well at its respective MOI. These samples were then serially diluted in 90 µL of LB broth. Following incubation of the dilution plates, colonies were counted in each dilution, and the bacterial concentration was computed as colony-forming units per milliliter (CFU/mL). These CFU/mL values were subsequently plotted graphically to illustrate the bacterial concentration dynamics over time at varying MOIs, enabling the comparison of bacterial reduction across different MOI values and time intervals (33).
Results
Isolation and Purification of Bacteriophage
Bacteriophages were successfully isolated from soil samples using the soft agar overlay method, and X. axonopodis pv. citri served as the host bacteria. The spot test indicated the presence of bacteriophages showing the clear plaques on the lawn of host bacteria. The isolated phage, specific to X. axonopodis pv. citri, produced several small circular plaques with a diameter ranging from 3-5 mm on the LB agar plates. These plaques were further analyzed to get uniform bacteriophage. A double-layer agar assay was performed for the purification of bacteriophages. The results are summarized in Table 1 and illustrated in Figure 1.
Table 1.
Results of the Spot Assay against Xanthomonas axonopodis pv. citri
|
S. No.
|
Bacteriophages
|
Area
|
Result
|
| 1 |
1 P |
Khanpur |
Positive ( + ) |
| 2 |
2 P |
Khanpur |
Positive ( + ) |
| 3 |
3 P |
Khanpur |
Positive ( + ) |
| 4 |
5 p |
Khanpur |
Positive ( + ) |
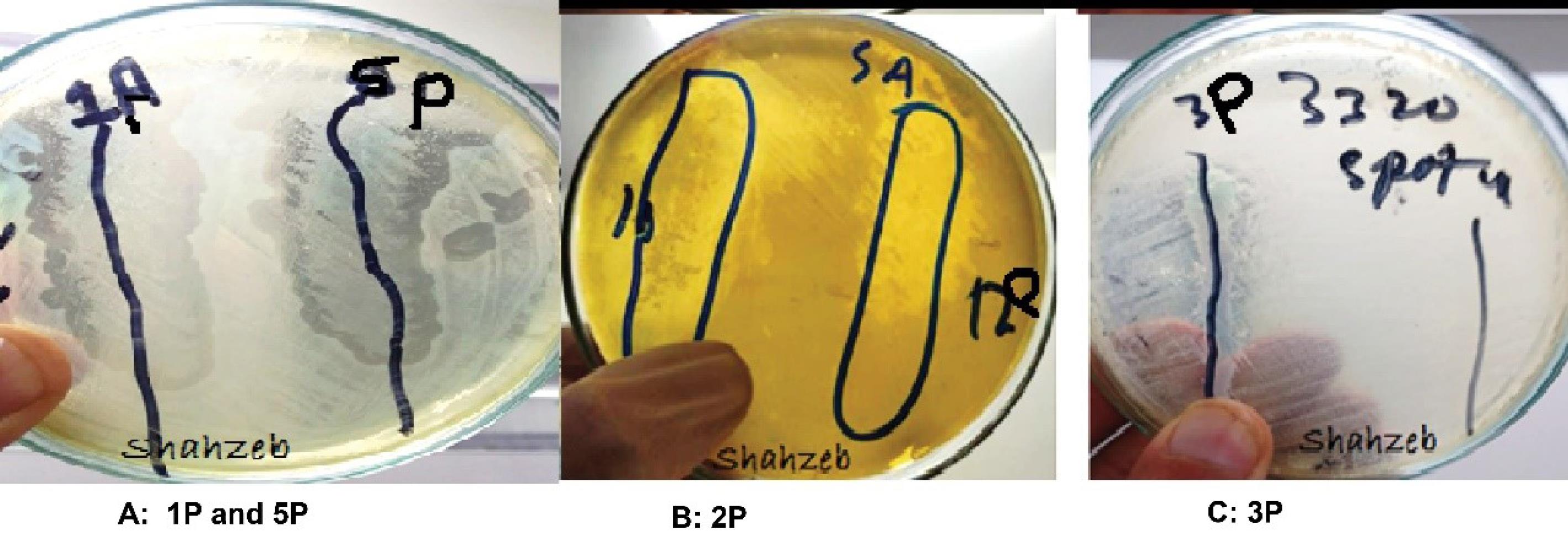
Figure 1.
Spot test confirming phage lysis on Xanthomonas axonopodis pv. citri
.
Spot test confirming phage lysis on Xanthomonas axonopodis pv. citri
Determination of Bacteriophage Titer
Purification and bacteriophage titer determination were done using a double-layer agar test. This approach enabled accurate quantification of plaque-forming units (PFU), confirming the presence and quantification of active bacteriophages. The PFUs are confirmed via Table 2 and Figure 2.
Table 2.
Result of Bacteriophage Titer Targeting Xanthomonas axonopodis pv. citri
|
S. No.
|
Bacteriophages
|
Titer (PFUs/mL)
|
| 1 |
1P |
3.0 × 108 |
| 2 |
2P |
2.7 × 108 |
| 3 |
3P |
3.5 × 109 |
| 4 |
5P |
4.0 × 109 |
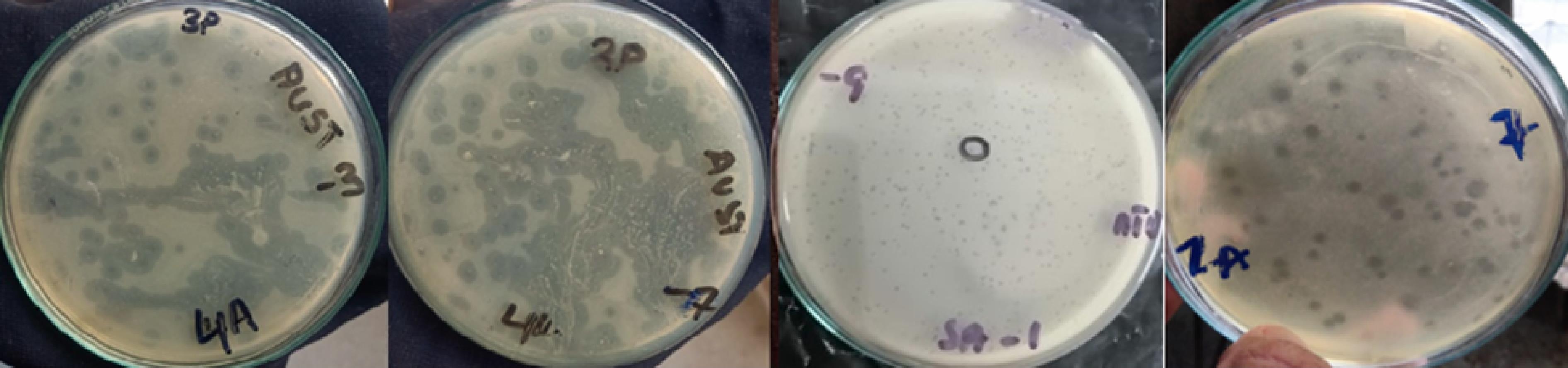
Figure 2.
Plaques of Different Titrations
.
Plaques of Different Titrations
Determination of Bacterial Growth Curve
Antibacterial activity was assessed using a bacterial growth reduction assay. The optical density method was used to develop a growth curve. Xanthomonas axonopodis pv. citri culture was inoculated in LB broth and optical density at 600 nm (OD600) was measured hourly to monitor bacterial growth dynamics. Simultaneously, aliquots of culture, prepared using the spreading technique, were plated on LB agar plates to measure the impact of bacterial proliferation. The recorded OD600 values are presented in Figure 3).
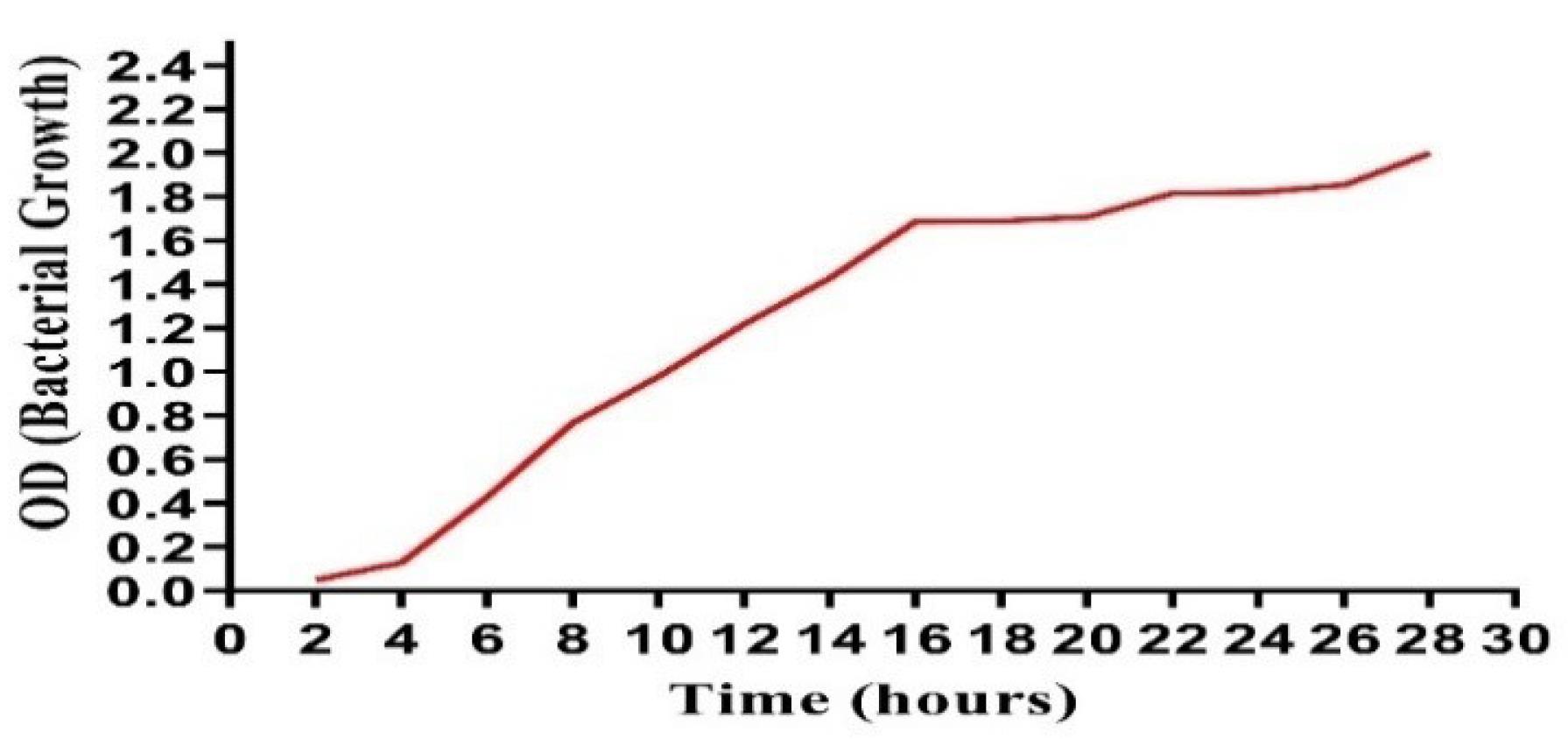
Figure 3.
Bacterial Growth Curve until 28 Hours
.
Bacterial Growth Curve until 28 Hours
Growth of Xanthomonas pv. citri at 7 °C
Xanthomonas axonopodis pv. citri was streaked on LB agar plates and incubated at 7 °C to assess the effect of temperature on its growth. The colonies developed a yellow pigment, characteristic of X. axonopodis, indicating active synthesis. These characteristics suggest that the bacteria continue to grow even at 7 °C and metabolize effectively, showing that low temperature does not affect the growth of X. axonopodis pv. citri (Figure 4).
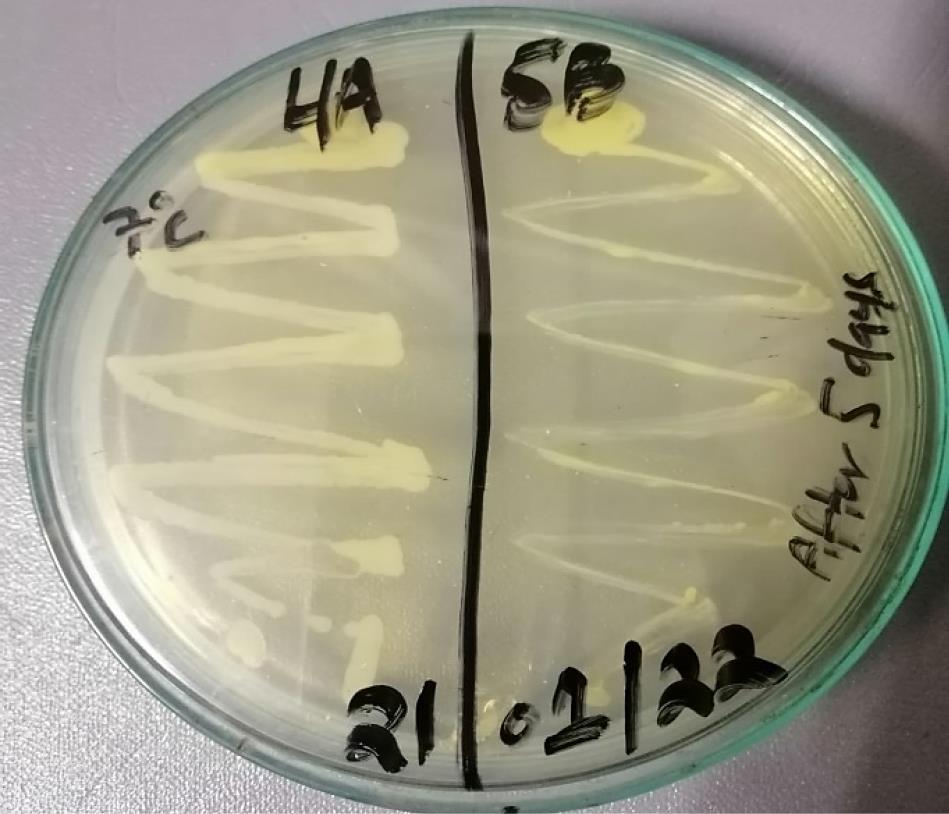
Figure 4.
Growth of Xanthomonas axonopodis pv. citri at 7 °C
.
Growth of Xanthomonas axonopodis pv. citri at 7 °C
The Activity of Isolated Bacteriophages at 7 °C against Xanthomonas axonopodis pv. citri
The activity of isolated bacteriophages specific to X. axonopodis pv. citri was observed at 7 °C using a double-layer technique. The phage produced several tiny asymmetrical elongated circular plaques on the LB Agar plates, indicating successful infection and lysis of bacterial cells. Our results revealed that phages retained their viability at low temperatures with an average of 201 plaques observed on each plate, suggesting that low temperature conditions did not affect the activity of bacteriophage (Figure 5).
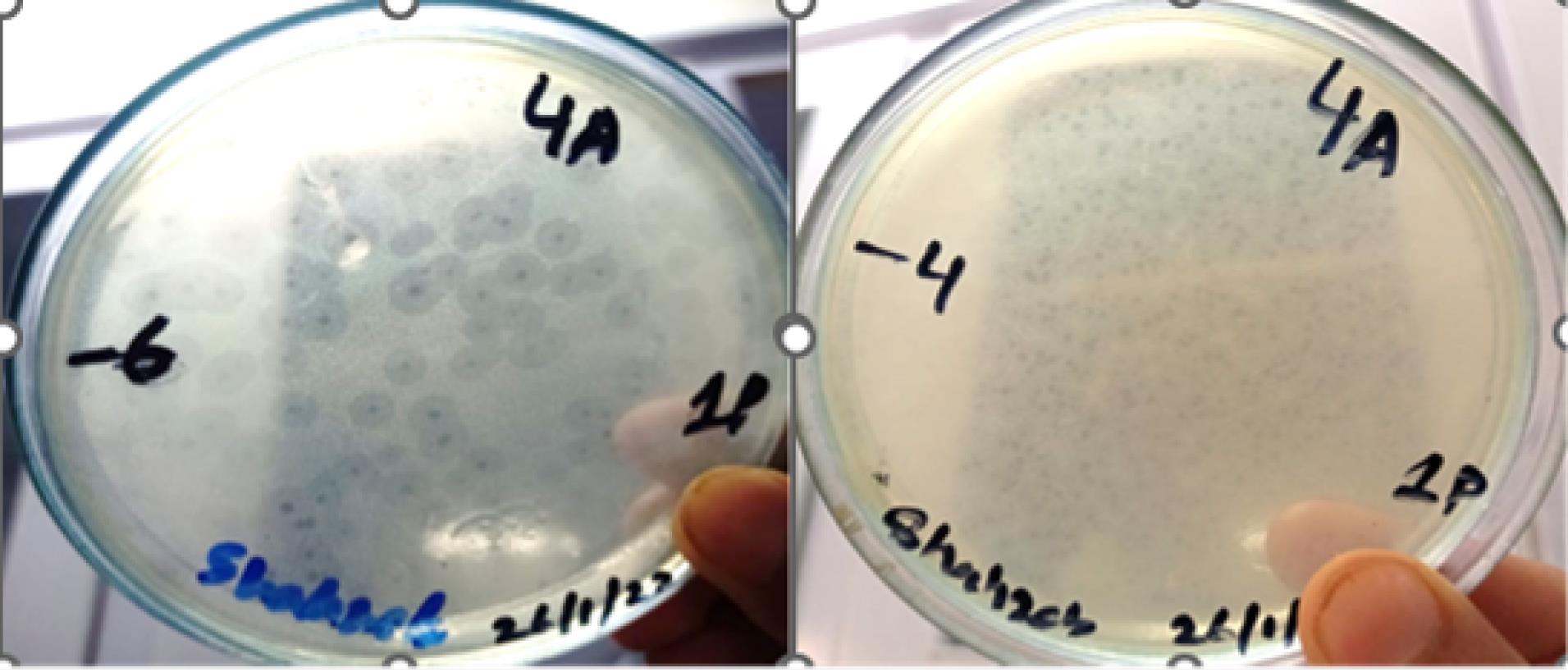
Figure 5.
Activity of Isolated Bacteriophages at 7 °C against X. axonopodis pv. citri
.
Activity of Isolated Bacteriophages at 7 °C against X. axonopodis pv. citri
Bacterial Growth Reduction by Bacteriophages
The isolated bacteriophages demonstrated a strong ability to reduce bacterial cell counts. To establish their lytic function, the phages were tested against X. axonopodis pv. citri in liquid medium using the bacterial reduction assay. During a 24-hour in vitro incubation period at 37 °C, efficient bacterial lysis by the selected bacteriophages was observed. The results of this phage-mediated bacterial reduction assay were compared to those of control samples. Compared to the controls, phage-infected samples exhibited a significant decrease in bacterial concentrations (Figure 6).
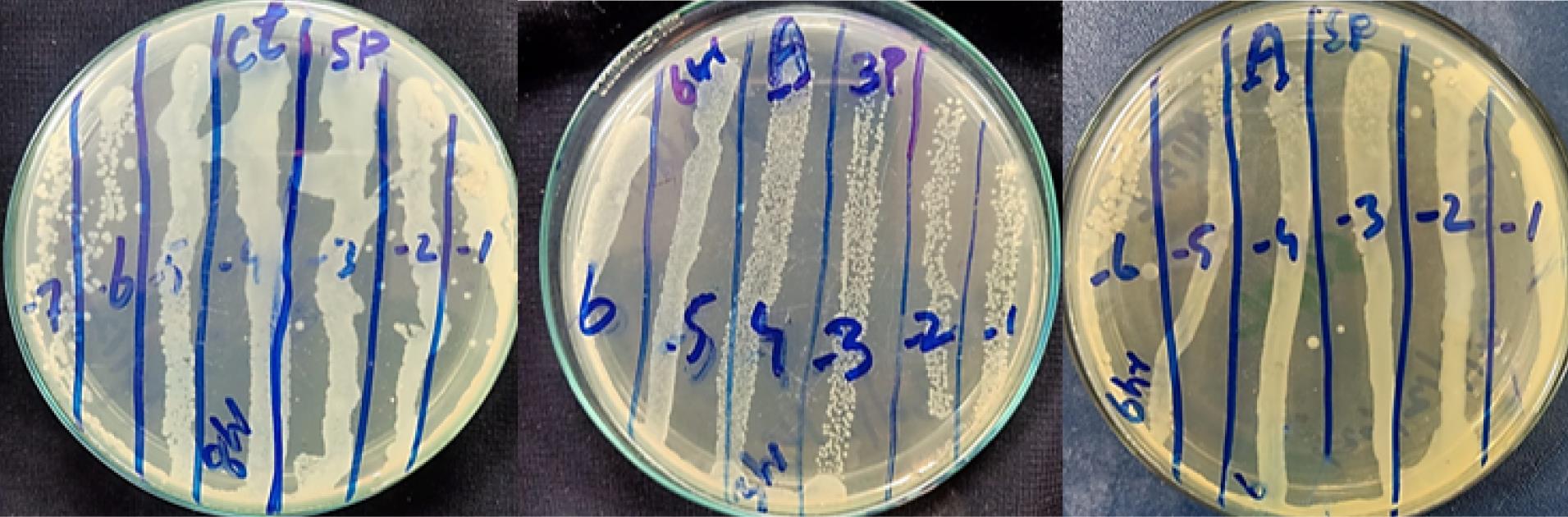
Figure 6.
In Vitro Effect of Bacteriophage on the Respective Bacteria: Reduction of Bacterial Cells by Corresponding Phages Compared with Control
.
In Vitro Effect of Bacteriophage on the Respective Bacteria: Reduction of Bacterial Cells by Corresponding Phages Compared with Control
Effect of Different Bacteriophages on Bacterial Growth at Different MOIs
The in vitro therapeutic efficacy of bacteriophage 1P against X. axonopodis pv. citri was evaluated using the bacterial growth reduction assay. Compared to control samples, phage infections resulted in a significant reduction in bacterial colonies. Bacteriophage 1P effectively inhibited X. axonopodis pv. citri growth for 24 hours at MOIs of 0.01, 10, and 100 (Figure 7). In contrast, at lower MOIs of 0.001 and 1, bacterial populations exhibited some resistance or regrowth after 20 hours. The considerable reduction in bacterial growth observed at MOI values of 0.01, 10, and 100 after 24 hours of incubation highlights the potential of phage 1P as a therapeutic agent against X. axonopodis pv. citri (Figure 7).
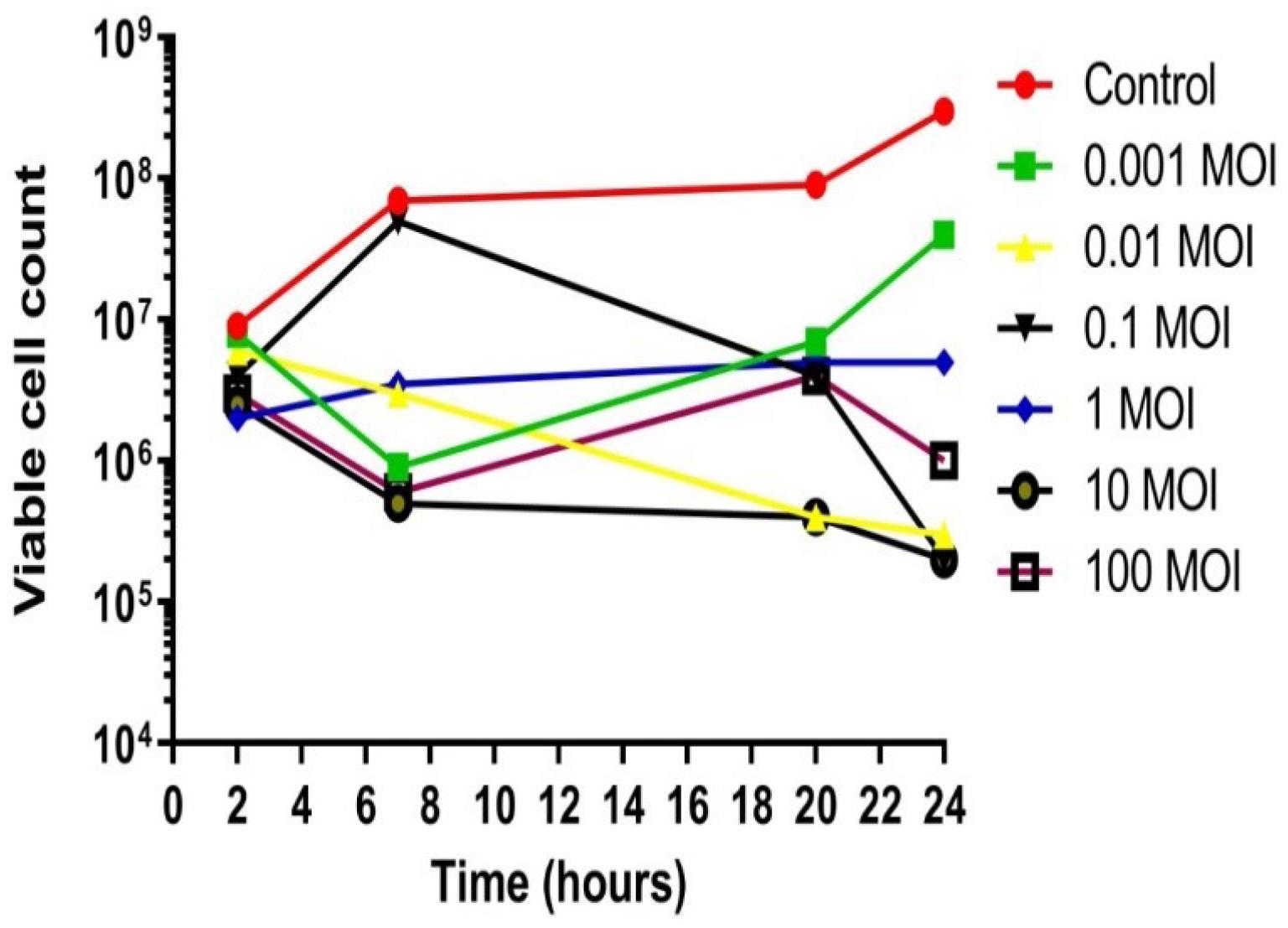
Figure 7.
In Vitro Effect of Phage 1P on Bacterial Growth: Reduction Compared to the Control. Viable cell count (PFUs/mL)
.
In Vitro Effect of Phage 1P on Bacterial Growth: Reduction Compared to the Control. Viable cell count (PFUs/mL)
Phage 2P demonstrated a decrease in bacterial viability across all tested MOIs (0.001, 0.01, 0.1, 1, and 100) during a 24-hour incubation (Figure 8). While a slight increase in bacterial resistance was noted at an MOI of 10 after 20 hours, phage 2P maintained maximum lytic activity between 20 and 24 hours. This sustained efficacy further indicates its therapeutic potential as an effective antibacterial agent (Figure 8).
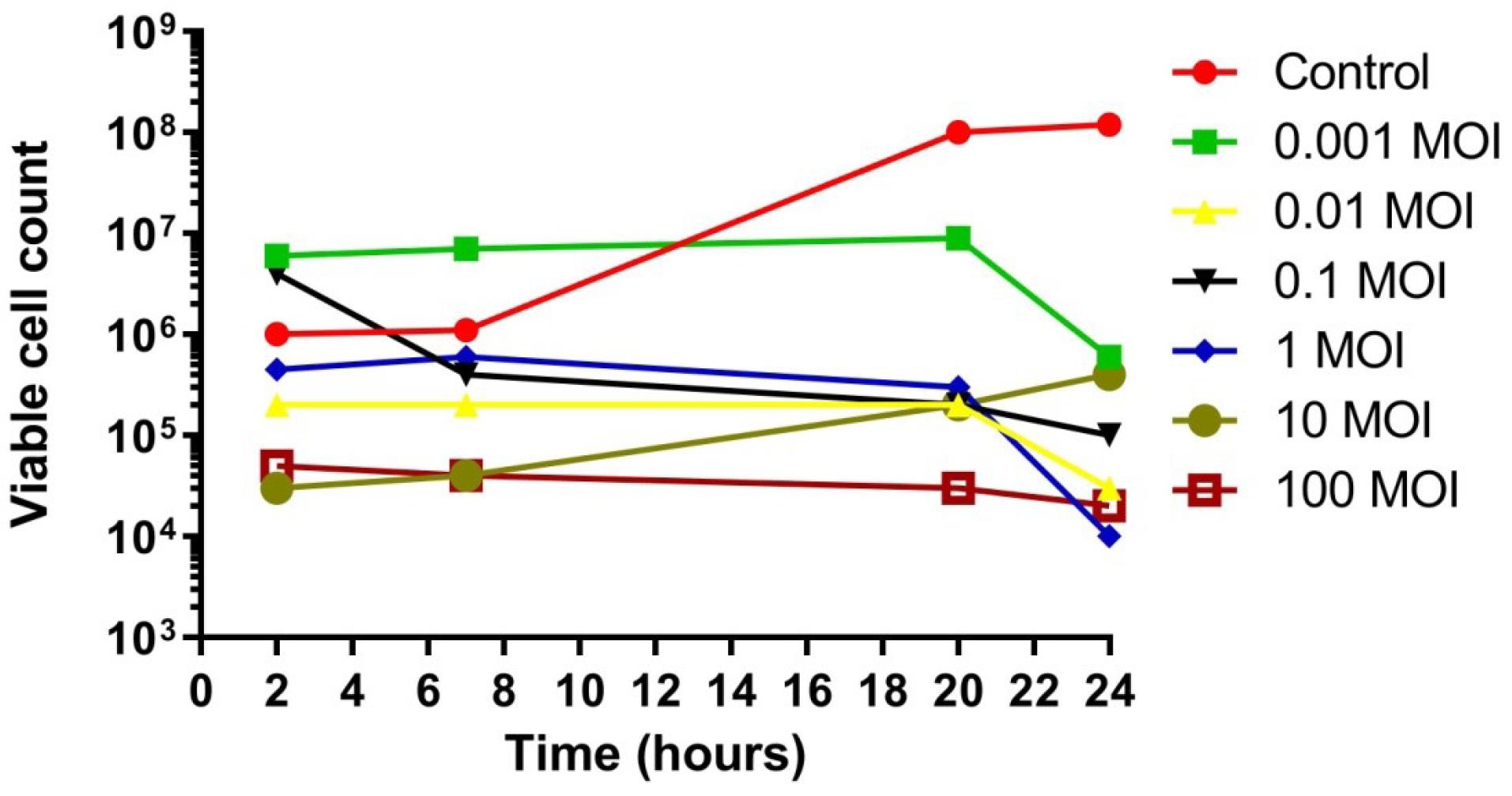
Figure 8.
In Vitro Effect of Bacteriophage 2P on Bacterial Growth: Reduction Compared to Control. Viable cell count (PFUs/mL)
.
In Vitro Effect of Bacteriophage 2P on Bacterial Growth: Reduction Compared to Control. Viable cell count (PFUs/mL)
Phage 3P demonstrated maximum lytic activity at 37 °C by effectively controlling the growth of X. axonopodis pv. citri. Phage 3P was found to effectively control bacterial growth until 24 hours. At MOIs of 0.01, 0.1, 1, 10, and 100, the bacteriophage 3P inhibited X. axonopodis pv. citri growth until 24 hours compared with the control, specifically at MOI 10, bacteriophages maintained bacterial suppression throughout the 24 hours, while at the lowest MOI (0.01), some bacterial resistance was observed after 16 hours. Overall, bacterial growth in the 3P-treated bacterial sample was greatly reduced, which also indicates the therapeutic potential of 3P (Figure 9).
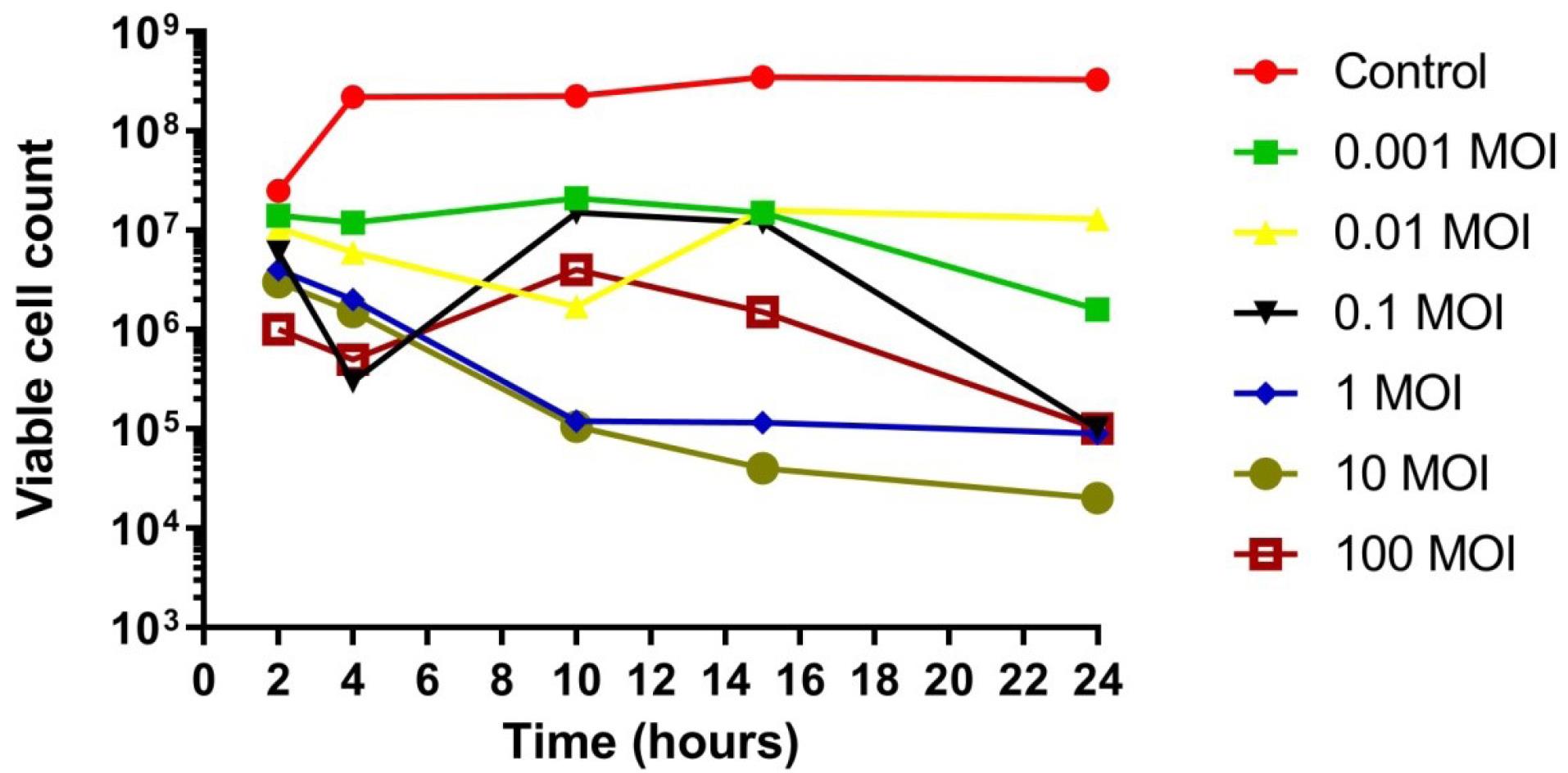
Figure 9.
In Vitro Effect of Bacteriophage 3P on Bacterial Growth: Reduction Compared to Control. Viable cell count (PFUs/mL)
.
In Vitro Effect of Bacteriophage 3P on Bacterial Growth: Reduction Compared to Control. Viable cell count (PFUs/mL)
Phage 5P served as a notable example of effective antibacterial action; infections by phage 5P led to a significant decrease in bacterial colonies compared to controls. Despite observing transient bacterial resistance during the initial two hours, this phage subsequently maintained inhibition of bacterial growth for the remainder of the 24-hour period, indicating promising potential for phage therapy (Figure 10). A general decline in bacterial populations was observed from 4 hours to 24 hours across all four tested phages. Nevertheless, transient increases in bacterial growth between 12 and 16 hours of incubation were attributed to the emergence of phage-resistant bacteria or the inactivation of phages.
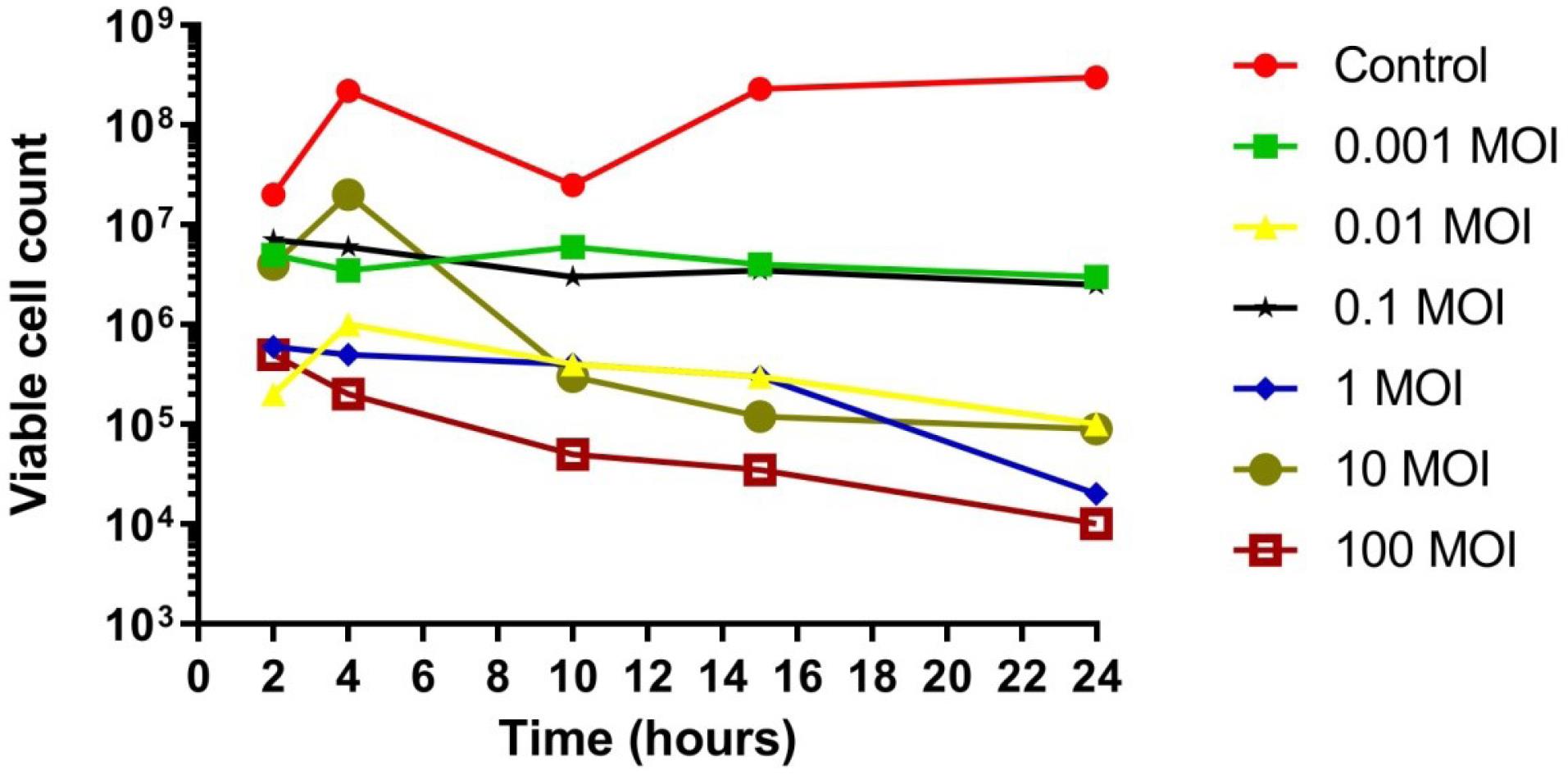
Figure 10.
In Vitro Effect of Bacteriophage 5P on Bacterial Growth: Reduction Compared to Control. Viable cell count (PFUs/mL)
.
In Vitro Effect of Bacteriophage 5P on Bacterial Growth: Reduction Compared to Control. Viable cell count (PFUs/mL)
Overall, this in vitro study demonstrated that bacteriophages isolated from soil samples possessed potent lytic effectiveness against X. axonopodis pv. citri, leading to a significant reduction in bacterial load. These findings suggest that these bacteriophages present a viable alternative to antibiotics for treating bacterial infections, positioning them as promising candidates for subsequent research and therapeutic development. These bacteriophages were thus selected for further investigation.
Discussion
AMR constitutes a formidable global health crisis, originating in the 20th century but now reaching alarmingly high levels worldwide, necessitating urgent intervention. Phage therapy offers a compelling strategy to combat this challenge. Economic impacts are substantial; an Organization for Economic Co-operation and Development (OECD) report indicates that high-income countries incur average annual costs of US10000 to US40000 for hospitalizations related to antibiotic-resistant bacterial infections. For middle-income and low-income countries, these costs approximate US12476 and US1025, respectively (39). Consequently, phage therapy has recently experienced a resurgence of interest (40). This work aims to contribute to this critical need by focusing on the isolation and characterization of effective phages. The therapeutic application of bacteriophages dates back approximately a century. Felix d’Herelle’s initial medical report on the use of bacteriophages to treat patients at the Hôpital des Enfants Malades in Paris was published in 1919 (41).
In this study, citrus fruits and soil served as primary sources for isolating X. axonopodis pv. citri-specific bacteriophages. Phage isolation and confirmation of lytic activity were performed using the spot test method. Four bacteriophages were subsequently selected for experiments and tested for their lytic activity and effectiveness against X. axonopodis pv. citri. Soil samples were initially collected from Khanpur region and transported to the laboratory for analysis. Of the 10 soil samples collected, only 4 yielded X. axonopodis pv. citri-specific bacteriophages, as detected by the spot assay technique. No lytic zones were observed in the remaining samples, even after enrichment.
Bacteriophages are obligate intracellular parasites, and their replication is entirely dependent on the growth and metabolism of their host bacteria (42). Consequently, phage recovery and amplification tend to be more successful in environments where the density of susceptible host bacteria is higher (43). Several studies suggest that the abundance of bacterial hosts directly influences the effectiveness of bacteriophages in altering microbial community composition (44,45). Since bacteriophages rely on specific bacterial hosts for replication (46), this observation can be attributed to variations in the concentration of X. axonopodis pv. citri at the sample collection sites, which could influence the availability of host bacteria for phage proliferation. When host bacteria are present in lower densities in the source soil, host-specific phages are less likely to have contact with each other. Most bacteriophages require a minimum bacterial density of approximately 103–104 CFU/mL to sustain successful multiplication (47).
Only clear plaques were chosen for further analysis based on plaque morphology, as clear zones indicate complete lysis of host bacteria X. axonopodis pv. citri. The isolated phage produced circular transparent plaques on double agar plates against the host bacteria. However, it was also observed that all the clear plagues were not the same size, which shows the presence of more than one phage in the soil. The plaque is made up of two circular layers, with a completely transparent inner center surrounded by a halo. The halo surrounding the plaque indicates that phage-produced soluble enzymes, such as depolymerase, degrade the bacterial cell walls (48). This observation is consistent with previous studies that have documented a wide range of plaque sizes even within a single phage collection, reflecting inherent genetic and functional diversity among phages (49). The presence of the depolymerase enzyme was shown by the halo, which diffused through the agar layer and broke down the bacterial capsular polysaccharide into distinct oligosaccharide components (50). Rahimzadeh et al achieved a titer of 20 × 108 PFU/mL using the double-layer agar assay (51). In an in vitro study, bacteriophages isolated from soil samples displayed lytic effectiveness against bacterial isolates from citrus fruits. These phages displayed lytic activity and successfully reduced the bacterial loads, indicating their potential to serve as a natural alternative to antibiotics. Various studies have been conducted to investigate antibacterial activity utilizing growth reduction assay against X. axonopodis pv. citri. For example, a study conducted on X. axonopodis pv. manihotis discussed methods for visualizing bacterial movement and growth in plants, emphasizing the relevance of growth dynamics in understanding pathogenicity (52). Another study focused on the growth dynamics of X. axonopodis pv. punicae using optical density measurements to evaluate the efficiency of antagonistic strains against this disease, stressing the importance of CFU counts in determining bacterial proliferation (53).
Our observations indicate a significant inhibitory effect of bacteriophages on the growth of X. axonopodis pv. citri, particularly within the initial 2 to 18 hours of treatment. At MOIs of both 0.001 and 1, we observed the emergence of some bacterial resistance to the bacteriophages after 20 hours. Specifically, X. axonopodis pv. citri populations were almost entirely suppressed until 18 hours. While a slight increase in bacterial count was observed after 20 hours, likely attributable to the emergence of phage-resistant mutants, this increase was still considerably lower compared to the control group. This phenomenon aligns with findings of Wang et al, who reported a continuous reduction in bacterial count over an 8-hour period with the novel bacteriophage vB_SauS_SA2, followed by the development of phage resistance (54).
Initially, bacteria showed some resistance to phage infection during the first two hours; however, phage activity could overcome this resistance effectively, suppressing bacterial growth up to 24 hours, which indicates promising potentialities of bacteriophage therapy. From 4 to 24 hours, all four tested phages showed a continuous decrease in bacterial populations. This delayed but effective lytic response aligns with previous reports suggesting that early-phase resistance may be due to host stress responses, adsorption inefficiency, or population heterogeneity (21). Our in vitro investigation revealed that bacteriophages isolated from soil exhibited potent lytic efficacy against citrus-associated bacteria, significantly reducing the bacterial load and underscoring their potential as alternatives to conventional antibiotics. A study by Lerdsittikul et al discovered a new bacteriophage, VL10, which demonstrated strong lytic activity against MRSA. Their results revealed a dramatic reduction in bacterial counts within the first 20 hours post-infection, consistent with our current findings. However, a subsequent resurgence in bacterial growth was observed after this period due to the development of resistance, which correlates with our own observation of higher bacterial counts after 20 hours (55).
Tuomala et al (2021) explored the application of phage therapy for the eradication of LA-MRSA in carrier pigs. They reported initial reductions in bacterial counts, but they also noted the evolution of resistance over time. These findings parallel our own observations regarding the emergence of resistance following sustained exposure at MOIs of 0.001 and 1. Such results suggest that phages have the potential to serve as an alternative to antibiotics for treating bacterial infections (16).
The outcomes of the current study are highly significant, as the isolated phage has the capability to lyse X. axonopodis pv. citri. By leveraging phage therapy as an alternative, this approach could pave the way for sustainable and targeted bacterial control in agricultural settings, ultimately enhancing crop protection while minimizing negative effects on beneficial microbes and the environment and serving as an important step toward phage therapy. This study resulted in the identification of X. axonopodis pv. citri -specific phage, which is a valued finding associated with microbiology and other fields.
Conclusion
A novel bacteriophage with high specificity for X. axonopodis pv. citri was successfully isolated and characterized. The isolated phage exhibited significant lytic activity, effectively inhibiting the growth of X. axonopodis pv. citri for over 24 hours. These results underscore the strong potential of this bacteriophage as a valuable biocontrol agent for citrus canker. It could be developed as an effective tool for disease management, providing both preventive and curative strategies against infections caused by X. axonopodis pv. citri.
Competing Interests
The authors declare that they have no competing interests.
Consent for Publication
Not applicable.
Data Availability Statement
The data will be made available when requested.
Ethical Approval
Not applicable.
Funding
The current research work was supported by a Research Grant provided by Higher Education Commission Pakistan through NRPU-HEC Project No. No: 10066/KPK/NRPU/R&D/HEC/2017.
References
- Read AF, Woods RJ. Antibiotic resistance management. Evol Med Public Health 2014; 2014(1):147. doi: 10.1093/emph/eou024 [Crossref] [ Google Scholar]
- Luby E, Ibekwe AM, Zilles J, Pruden A. Molecular methods for assessment of antibiotic resistance in agricultural ecosystems: prospects and challenges. J Environ Qual 2016; 45(2):441-53. doi: 10.2134/jeq2015.07.0367 [Crossref] [ Google Scholar]
- Ghosh S, LaPara TM. The effects of subtherapeutic antibiotic use in farm animals on the proliferation and persistence of antibiotic resistance among soil bacteria. ISME J 2007; 1(3):191-203. doi: 10.1038/ismej.2007.31 [Crossref] [ Google Scholar]
- Haghighatpanah M, Mozaffari Nejad AS, Mojtahedi A, Amirmozafari N, Zeighami H. Detection of extended-spectrum β-lactamase (ESBL) and plasmid-borne bla(CTX-M) and bla(TEM) genes among clinical strains of Escherichia coli isolated from patients in the north of Iran. J Glob Antimicrob Resist 2016; 7:110-3. doi: 10.1016/j.jgar.2016.08.005 [Crossref] [ Google Scholar]
- Tabassum R, Alvi IA, Asif M, Basit A, Rehman SU. TSP, a virulent podovirus can control the growth of Staphylococcus aureus till 12 hours. Res Sq [Preprint]. February 18, 2021. doi: 10.21203/rs.3.rs-170860/v1.
- Karami P, Bazmamoun H, Sedighi I, Mozaffari Nejad AS, Aslani MM, Alikhani MY. Antibacterial resistance patterns of extended spectrum β-lactamase -producing enteropathogenic Escherichia coli strains isolated from children. Arab J Gastroenterol 2017; 18(4):206-9. doi: 10.1016/j.ajg.2017.11.004 [Crossref] [ Google Scholar]
- Hoseini N, Sedighi I, Mozaffari Nejad AS, Alikhani MY. Phenotypic and genotypic detection of AmpC enzymes in clinical isolates of Escherichia coli and Klebsiella pneumoniae. J Krishna Inst Med Sci Univ 2017;6(3)10-8.
- Haghighatpanah M, Zeighami H, Mozaffari Nejad AS, Hajipour N. Phylogenetic groups and antimicrobial resistance characteristics of Escherichia coli strains isolated from clinical samples in north Iran. Arab J Gastroenterol 2022; 23(2):102-7. doi: 10.1016/j.ajg.2022.02.003 [Crossref] [ Google Scholar]
- Sedighi I, Mozaffari Nejad AS, Amanati A, Nakhaei S, Alikhani MY. Virulence factors and antibiotic resistance in uropathogenic and commensal Escherichia coli isolates. J Krishna Inst Med Sci Univ 2016; 5(4):50-7. [ Google Scholar]
- Markwart R, Saito H, Harder T, Tomczyk S, Cassini A, Fleischmann-Struzek C. Epidemiology and burden of sepsis acquired in hospitals and intensive care units: a systematic review and meta-analysis. Intensive Care Med 2020; 46(8):1536-51. doi: 10.1007/s00134-020-06106-2 [Crossref] [ Google Scholar]
- Monegro AF, Muppidi V, Regunath H. Hospital-acquired infections. In: StatPearls [Internet]. Treasure Island, FL: StatPearls Publishing; 2025.
- Manyi-Loh C, Mamphweli S, Meyer E, Okoh A. Antibiotic use in agriculture and its consequential resistance in environmental sources: potential public health implications. Molecules 2018; 23(4):795. doi: 10.3390/molecules23040795 [Crossref] [ Google Scholar]
- Mulchandani R, Wang Y, Gilbert M, Van Boeckel TP. Global trends in antimicrobial use in food-producing animals: 2020 to 2030. PLOS Glob Public Health 2023; 3(2):e0001305. doi: 10.1371/journal.pgph.0001305 [Crossref] [ Google Scholar]
- Mann A, Nehra K, Rana JS, Dahiya T. Antibiotic resistance in agriculture: perspectives on upcoming strategies to overcome upsurge in resistance. Curr Res Microb Sci 2021; 2:100030. doi: 10.1016/j.crmicr.2021.100030 [Crossref] [ Google Scholar]
- Balogh B, Canteros BI, Stall RE, Jones JB. Control of citrus canker and citrus bacterial spot with bacteriophages. Plant Dis 2008; 92(7):1048-52. doi: 10.1094/pdis-92-7-1048 [Crossref] [ Google Scholar]
- Tuomala H, Verkola M, Meller A, Van der Auwera J, Patpatia S, Järvinen A. Phage treatment trial to eradicate LA-MRSA from healthy carrier pigs. Viruses 2021; 13(10):1888. doi: 10.3390/v13101888 [Crossref] [ Google Scholar]
- Azizian R, Nasser A, Askari H, Taheri Kalani M, Sadeghifard N, Pakzad I. Sewage as a rich source of phage study against Pseudomonas aeruginosa PAO. Biologicals 2015; 43(4):238-41. doi: 10.1016/j.biologicals.2015.05.004 [Crossref] [ Google Scholar]
- Chanishvili N. Phage therapy--history from Twort and d’Herelle through Soviet experience to current approaches. Adv Virus Res 2012; 83:3-40. doi: 10.1016/b978-0-12-394438-2.00001-3 [Crossref] [ Google Scholar]
- Lin TL, Hsieh PF, Huang YT, Lee WC, Tsai YT, Su PA. Isolation of a bacteriophage and its depolymerase specific for K1 capsule of Klebsiella pneumoniae: implication in typing and treatment. J Infect Dis 2014; 210(11):1734-44. doi: 10.1093/infdis/jiu332 [Crossref] [ Google Scholar]
- Henry M, Lavigne R, Debarbieux L. Predicting in vivo efficacy of therapeutic bacteriophages used to treat pulmonary infections. Antimicrob Agents Chemother 2013; 57(12):5961-8. doi: 10.1128/aac.01596-13 [Crossref] [ Google Scholar]
- Abedon ST, Kuhl SJ, Blasdel BG, Kutter EM. Phage treatment of human infections. Bacteriophage 2011; 1(2):66-85. doi: 10.4161/bact.1.2.15845 [Crossref] [ Google Scholar]
- Abedon ST. Phage therapy of pulmonary infections. Bacteriophage 2015; 5(1):e1020260. doi: 10.1080/21597081.2015.1020260 [Crossref] [ Google Scholar]
- Chang RY, Wallin M, Lin Y, Leung SS, Wang H, Morales S. Phage therapy for respiratory infections. Adv Drug Deliv Rev 2018; 133:76-86. doi: 10.1016/j.addr.2018.08.001 [Crossref] [ Google Scholar]
- Hoyle N, Zhvaniya P, Balarjishvili N, Bolkvadze D, Nadareishvili L, Nizharadze D. Phage therapy against Achromobacterxylosoxidans lung infection in a patient with cystic fibrosis: a case report. Res Microbiol 2018; 169(9):540-2. doi: 10.1016/j.resmic.2018.05.001 [Crossref] [ Google Scholar]
- Aslam S, Courtwright AM, Koval C, Lehman SM, Morales S, Furr CL. Early clinical experience of bacteriophage therapy in 3 lung transplant recipients. Am J Transplant 2019; 19(9):2631-9. doi: 10.1111/ajt.15503 [Crossref] [ Google Scholar]
- Wright A, Hawkins CH, Anggård EE, Harper DR. A controlled clinical trial of a therapeutic bacteriophage preparation in chronic otitis due to antibiotic-resistant Pseudomonas aeruginosa; a preliminary report of efficacy. Clin Otolaryngol 2009; 34(4):349-57. doi: 10.1111/j.1749-4486.2009.01973.x [Crossref] [ Google Scholar]
- Dedrick RM, Guerrero-Bustamante CA, Garlena RA, Russell DA, Ford K, Harris K. Engineered bacteriophages for treatment of a patient with a disseminated drug-resistant Mycobacterium abscessus. Nat Med 2019; 25(5):730-3. doi: 10.1038/s41591-019-0437-z [Crossref] [ Google Scholar]
- Chaudhry WN, Concepción-Acevedo J, Park T, Andleeb S, Bull JJ, Levin BR. Synergy and order effects of antibiotics and phages in killing Pseudomonas aeruginosa biofilms. PLoS One 2017; 12(1):e0168615. doi: 10.1371/journal.pone.0168615 [Crossref] [ Google Scholar]
- Henriksen K, Rørbo N, Rybtke ML, Martinet MG, Tolker-Nielsen T, Høiby N. P aeruginosa flow-cell biofilms are enhanced by repeated phage treatments but can be eradicated by phage–ciprofloxacin combination: monitoring the phage–P aeruginosa biofilms interactions. Pathog Dis 2019; 77(2):ftz011. doi: 10.1093/femspd/ftz011 [Crossref] [ Google Scholar]
- Milho C, Andrade M, Vilas Boas D, Alves D, Sillankorva S. Antimicrobial assessment of phage therapy using a porcine model of biofilm infection. Int J Pharm 2019; 557:112-23. doi: 10.1016/j.ijpharm.2018.12.004 [Crossref] [ Google Scholar]
- Yang Q, Le S, Zhu T, Wu N. Regulations of phage therapy across the world. Front Microbiol 2023; 14:1250848. doi: 10.3389/fmicb.2023.1250848 [Crossref] [ Google Scholar]
- Bibi Z, Abbas Z, Rehman SU. The phage PE1 isolated from hospital sewage reduces the growth of Escherichia coli. Biocontrol Sci Technol 2016; 26(2):181-8. doi: 10.1080/09583157.2015.1086311 [Crossref] [ Google Scholar]
- Alvi IA, Asif M, Tabassum R, Aslam R, Abbas Z, Rehman SU. RLP, a bacteriophage of the family Podoviridae, rescues mice from bacteremia caused by multi-drug-resistant Pseudomonas aeruginosa. Arch Virol 2020; 165(6):1289-97. doi: 10.1007/s00705-020-04601-x [Crossref] [ Google Scholar]
- O’Flaherty S, Ross RP, Meaney W, Fitzgerald GF, Elbreki MF, Coffey A. Potential of the polyvalent anti-Staphylococcus bacteriophage K for control of antibiotic-resistant staphylococci from hospitals. Appl Environ Microbiol 2005; 71(4):1836-42. doi: 10.1128/aem.71.4.1836-1842.2005 [Crossref] [ Google Scholar]
- Sunagar R, Patil SA, Chandrakanth RK. Bacteriophage therapy for Staphylococcus aureus bacteremia in streptozotocin-induced diabetic mice. Res Microbiol 2010; 161(10):854-60. doi: 10.1016/j.resmic.2010.09.011 [Crossref] [ Google Scholar]
- Brenner S, Streisinger G, Horne RW, Champe SP, Barnett L, Benzer S. Structural components of bacteriophage. J Mol Biol 1959; 1(3):281-92. doi: 10.1016/s0022-2836(59)80035-8 [Crossref] [ Google Scholar]
- Cuevas DA, Edwards RA. PMAnalyzer: a new web interface for bacterial growth curve analysis. Bioinformatics 2017; 33(12):1905-6. doi: 10.1093/bioinformatics/btx084 [Crossref] [ Google Scholar]
- Abedon ST. Bacteriophage clinical use as antibacterial “drugs”: utility and precedent. Microbiol Spectr 2017;5(4):10.1128/microbiolspec.bad-0003-2016. doi: 10.1128/microbiolspec.BAD-0003-2016.
- Adamie BA, Akwar HT, Arroyo M, Bayko H, Hafner M, Harrison S, et al. Forecasting the Fallout from AMR: Economic Impacts of Antimicrobial Resistance in Food-Producing Animals. World Organisation for Animal Health; 2024.
- Sahoo K, Meshram S. The evolution of phage therapy: a comprehensive review of current applications and future innovations. Cureus 2024; 16(9):e70414. doi: 10.7759/cureus.70414 [Crossref] [ Google Scholar]
- Lin DM, Koskella B, Lin HC. Phage therapy: an alternative to antibiotics in the age of multi-drug resistance. World J Gastrointest Pharmacol Ther 2017; 8(3):162-73. doi: 10.4292/wjgpt.v8.i3.162 [Crossref] [ Google Scholar]
- Naureen Z, Dautaj A, Anpilogov K, Camilleri G, Dhuli K, Tanzi B. Bacteriophages presence in nature and their role in the natural selection of bacterial populations. Acta Biomed 2020; 91(13-S):e2020024. doi: 10.23750/abm.v91i13-S.10819 [Crossref] [ Google Scholar]
- Ranveer SA, Dasriya V, Ahmad MF, Dhillon HS, Samtiya M, Shama E. Positive and negative aspects of bacteriophages and their immense role in the food chain. NPJ Sci Food 2024; 8(1):1. doi: 10.1038/s41538-023-00245-8 [Crossref] [ Google Scholar]
- Gundersen MS, Fiedler AW, Bakke I, Vadstein O. The impact of phage treatment on bacterial community structure is minor compared to antibiotics. Sci Rep 2023; 13(1):21032. doi: 10.1038/s41598-023-48434-5 [Crossref] [ Google Scholar]
- Castledine M, Buckling A. Critically evaluating the relative importance of phage in shaping microbial community composition. Trends Microbiol 2024; 32(10):957-69. doi: 10.1016/j.tim.2024.02.014 [Crossref] [ Google Scholar]
- Kasman LM, Porter LD. Bacteriophages. In: StatPearls [Internet]. Treasure Island, FL: StatPearls Publishing; 2025.
- Wiggins BA, Alexander M. Minimum bacterial density for bacteriophage replication: implications for significance of bacteriophages in natural ecosystems. Appl Environ Microbiol 1985; 49(1):19-23. doi: 10.1128/aem.49.1.19-23.1985 [Crossref] [ Google Scholar]
- Hughes KA, Sutherland IW, Jones MV. Biofilm susceptibility to bacteriophage attack: the role of phage-borne polysaccharide depolymerase. Microbiology (Reading) 1998; 144(Pt 11):3039-47. doi: 10.1099/00221287-144-11-3039 [Crossref] [ Google Scholar]
- Ceyssens PJ, Miroshnikov K, Mattheus W, Krylov V, Robben J, Noben JP. Comparative analysis of the widespread and conserved PB1-like viruses infecting Pseudomonas aeruginosa. Environ Microbiol 2009; 11(11):2874-83. doi: 10.1111/j.1462-2920.2009.02030.x [Crossref] [ Google Scholar]
- Adams MH, Park BH. An enzyme produced by a phage-host cell system II The properties of the polysaccharide depolymerase. Virology 1956; 2(6):719-36. doi: 10.1016/0042-6822(56)90054-x [Crossref] [ Google Scholar]
- Rahimzadeh G, Resch G, Rezai MS, Nemati Hevelaee E. Characterization and lytic activity of isolated Escherichia coli bacteriophages against Escherichia coli in vitro. Iran J Med Sci 2020; 45(4):298-303. doi: 10.30476/ijms.2019.45420 [Crossref] [ Google Scholar]
- Cohn M, Shybut M, Dahlbeck D, Staskawicz B. Assays to assess virulence of Xanthomonas axonopodis pv manihotis on cassava. Bio Protoc 2015; 5(13):e1522. doi: 10.21769/BioProtoc.1522 [Crossref] [ Google Scholar]
- Giri MS, Rabinal CA, Raju J, Jayalakshmi K. Isolation and screening of anti-microbial compounds produced by fluorescent pseudomonads against Xanthomonas axonopodis pv punicae. J Plant Dis Sci 2023; 18(2):109-14. doi: 10.48165/jpds.2023.1802.08 [Crossref] [ Google Scholar]
- Wang J, Zhao F, Sun H, Wang Q, Zhang C, Liu W. Isolation and characterization of the Staphylococcus aureus bacteriophage vB_SauS_SA2. AIMS Microbiol 2019; 5(3):285-307. doi: 10.3934/microbiol.2019.3.285 [Crossref] [ Google Scholar]
- Lerdsittikul V, Apiratwarrasakul S, Atithep T, Withatanung P, Indrawattana N, Pumirat P. Isolation and characterisation of a novel Silviavirus bacteriophage promising antimicrobial agent against methicillin-resistant Staphylococcus aureus infections. Sci Rep 2024; 14(1):9251. doi: 10.1038/s41598-024-59903-w [Crossref] [ Google Scholar]