Avicenna Journal of Clinical Microbiology and Infection. 12(3):152-162.
doi: 10.34172/ajcmi.3604
Review Article
The Role of Crucial Genes in the Multiplication Cycle, Immune Evasion, and Development of Novel Treatments in Intracellular Bacteria Chlamydia: A Focus on Chlamydia trachomatis
Mohammad Ebrahim Karimbakhsh Conceptualization, Data curation, Investigation, Resources, Software, Visualization, Writing – original draft, Writing – review & editing, 1 
Rouzbeh Sojoudi Masuleh Data curation, Writing – original draft, 2
Alireza Kazemi Visualization, 3
Hamid Reza Goli Formal analysis, Software, Writing – review & editing, 1
Mehrdad Gholami Methodology, Project administration, Software, Supervision, Validation, 1, *
Author information:
1Department of Medical Microbiology and Virology, Faculty of Medicine, Mazandaran University of Medical Sciences, Sari, Iran
2Department of Immunology, Faculty of Medical Sciences, Tarbiat Modares University, Tehran, Iran
3Department of Cell and Molecular Biology, Faculty of Life Sciences, Shahr-e-Qods Branch, Islamic Azad University, Tehran, Iran
Abstract
Background: Chlamydia species are known to cause significant health challenges in humans, including respiratory tract infections and sexually transmitted diseases. Among them, Chlamydia trachomatis is one of the most prevalent sexually transmitted pathogens worldwide. Although C. trachomatis exhibits minimal documented antibiotic resistance, the development of advanced therapeutic strategies, such as peptide-based drugs and vaccines, presents promising avenues for mitigating resistance and enhancing treatment outcomes.
Methods: This study investigated the role of key genes involved in Chlamydia biology, including TNT, FtsK, Euo, and ClpX, through a comprehensive review of scientific literature from reputable databases, such as Google Scholar. Diagnostic methods, such as polymerase chain reaction, ligase chain reaction, and nucleic acid sequence-based amplification, were highlighted for their accuracy in detecting pathogenic factors, such as the type III secretion system.
Results: The analysis demonstrated that Chlamydia possesses sophisticated immune evasion mechanisms by targeting neutrophils, dendritic cells (DCs), and macrophages. The reproductive cycle of C. trachomatis and its genetic components were found to be intricately linked to its pathogenicity. Furthermore, understanding immune evasion strategies and key genes related to the bacterial lifecycle provides valuable insights into disease progression and potential therapeutic targets.
Conclusion: The findings of this study highlight the significance of unraveling the complex biology of Chlamydia species. A comprehensive understanding of its genetic makeup, immune evasion strategies, and pathogenic mechanisms is essential for developing novel preventive measures, effective therapies, and accurate diagnostic techniques. Future research in these areas is crucial for mitigating the public health impact of Chlamydia infections.
Keywords: Chlamydia trachomatis, Immune evasion, Antibiotic therapy, Virulence gene
Copyright and License Information
© 2025 The Author(s); Published by Hamadan University of Medical Sciences.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (
https://creativecommons.org/licenses/by/4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Please cite this article as follows: Karimbakhsh ME, Sojoudi Masuleh R, Kazemi A, Goli H, Gholami M. The role of crucial genes in the multiplication cycle, immune evasion, and development of novel treatments in intracellular bacteria chlamydia: a focus on chlamydia trachomatis. Avicenna J Clin Microbiol Infect. 2025;12(3):152-162. doi:10.34172/ajcmi.3604
Introduction
Since its discovery in 1907, Chlamydia has been extensively characterized, leading to significant advancements in understanding its biology and pathogenicity. This evolution has led to the development of various infections, including lung infections (e.g., Chlamydia pneumoniae), sexually transmitted infections (STIs), and abortions. However, due to the remarkable adaptability of this bacterium, novel genetic variants have emerged over time. While adaptability presents challenges in treatment and control, it also underscores the organism’s resilience and biological complexity (1,2). Chlamydia exhibits a tropism for various epithelial tissues, leading to diverse clinical manifestations (3). The biphasic developmental cycle of Chlamydia involves two distinct forms, namely, elementary bodies (EB) and reticulate bodies (RB). Chlamydia relies on eukaryotic host cells for survival and replication (4). The Chlamydia genus includes medically significant species, such as C. trachomatis, which is a major human pathogen (5). Additionally, other species in this family (C. abortus, C. caviae, C. felis, and C. psittaci) cause zoonotic infections (6). Chlamydia commonly leads to asymptomatic infections in both men and women. These infections may go undiagnosed and prevent treatment with appropriate medications, resulting in substantial tissue damage (7). C. trachomatis can cause various infections and is associated with several health concerns, including epididymitis in men and fertility problems in women (8). Additionally, vertical transmission from the mother to the child can result in severe symptoms, underscoring the importance of early diagnosis and prompt treatment. Routine screening for asymptomatic Chlamydia infections is crucial for preventing complications and limiting transmission within high-risk populations (9,10). Other bacterial complications include ectopic pregnancy and pelvic inflammatory disease (1,2). Chronic Chlamydia infections can result in irreversible tissue damage and sustained inflammation in affected tissues (11).
The genetic composition of the outer membrane of C. trachomatis, which affects various tissues and induces diverse symptoms, divides C. trachomatis into distinct serological variants. Serotypes A, B, and C primarily infect ocular epithelial cells, leading to trachoma, a major cause of blindness worldwide (12). Genital infections caused by C. trachomatis, primarily associated with serotypes D–K, represent the most common bacterial STIs worldwide (13). These serotypes lead to conjunctivitis, urinary tract infections, and pneumonia in children, as well as urogenital and extragenital infections in adults (14,15). Serotypes L1, L2, and L3 of C. trachomatis are responsible for lymphogranuloma venereum, an STI characterized by invasive infection of the lymphatic system (16). A summary of this information is displayed in Figure 1.
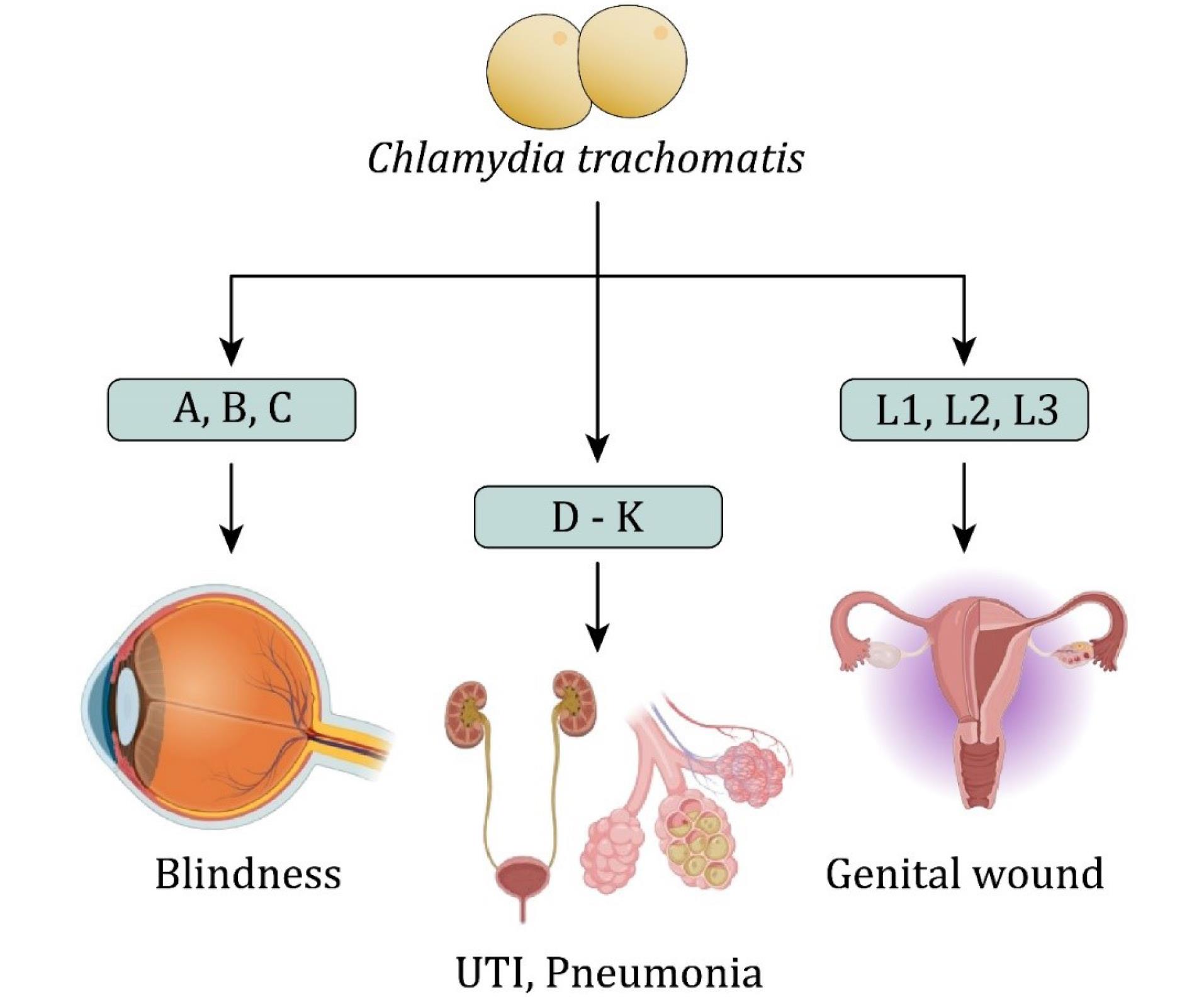
Figure 1.
Chlamydia trachomatis Serovars Associated With Distinct Human Infections. Note. UTI: Unitary tract infection. Serovars A, B, and C are etiological agents of trachoma, which can result in visual impairments. In addition, serovars D–K are implicated in urogenital infections, including UTIs and pneumonia. Moreover, serovars L1, L2, and L3 are causative agents of lymphogranuloma venereum, characterized by genital ulceration and lymphatic involvement.
.
Chlamydia trachomatis Serovars Associated With Distinct Human Infections. Note. UTI: Unitary tract infection. Serovars A, B, and C are etiological agents of trachoma, which can result in visual impairments. In addition, serovars D–K are implicated in urogenital infections, including UTIs and pneumonia. Moreover, serovars L1, L2, and L3 are causative agents of lymphogranuloma venereum, characterized by genital ulceration and lymphatic involvement.
Studying C. trachomatis through immunology, epidemiology, and bioinformatics facilitates the recognition of important processes, elucidates its infection cycles, and highlights complex host-pathogen interactions. Moreover, C. trachomatis exhibits a distinct dependence on specific host species and tissue tropism. This characteristic underscores its role in infertility, host immune responses, and vaccine development against human infections (17,18). In 2024, Poston reviewed recent advancements in vaccine development and protective adjuvants. These findings are critical for enhancing vaccine efficacy, and the proposed strategies hold potential for future research applications (19). Additionally, Chiarelli et al demonstrated that C. trachomatis undergoes a complex developmental cycle with three distinct cell types (EB, IB, and RB). Asymmetric division suggests that RBs exhibit stem cell-like properties, generating IBs as a renewable intermediate stage during each replication cycle (20).
Despite significant advancements in our understanding of the Chlamydia life cycle and immune evasion mechanisms, critical gaps persist in the identification of specific targets for therapeutic interventions. Bridging these gaps will facilitate the development of novel diagnostic and therapeutic strategies. This review explores the diverse immune evasion strategies employed by Chlamydia spp., the virulence-associated genes contributing to pathogenicity, and the most effective detection and treatment approaches.
Structure, Cycle of Growth, and Persistent Infection in Chlamydia
Chlamydia spp. exhibit a distinct two-phase developmental cycle for persistence and propagation, consisting of EB and RB, each fulfilling a specific function. In this cycle, the Euo regulatory protein plays a pivotal role in facilitating the transition to the EB form. The progression of the cycle is regulated by Euo expression levels, although the primary formation of RB remains unaffected (20-22). The EB form does not proliferate; rather, it enters host cells approximately eight hours post-infection and rapidly transforms into the RB form. Approximately twenty hours post-infection, RBs replicate via binary fission; subsequently, they transit into EBs, leading to an increased EB population upon host cell lysis (23-25). Given the absence of a homologous FtsZ protein and the lack of peptidoglycan recognition, investigations have been conducted to identify alternative essential proteins; (1) BacACT, an analogous of bactophilin, plays a significant role in preserving cellular morphology, as its reduced expression may lead to structural alterations in Chlamydia (26,27). (2) The Chlamydia proteins MreB and Pbp2 are crucial for cell division. Pbp2, a high-molecular-weight penicillin-binding protein, is essential for bacterial cell proliferation, whereas MreB regulates the maintenance of Chlamydia’s rod shape. The interaction among MreB, Pbp2, and Ftsk suggests their cooperative role in Chlamydia cell division. During bacterial proliferation, Ftsk is required for the proper segregation and formation of RBs and EBs. The findings suggest that MreB may functionally compensate for the absence of FtsZ during Chlamydia cell division (27,28). (3) Among the two key proteins regulating Chlamydia survival and developmental cycle, ClpX functions as an adenosine triphosphate-dependent protease (29).
Tunnel nanotubes (TNTs) facilitate the spread of Chlamydia from infected cells. Nanotubes are transient cellular structures that enable material exchange and intercellular communication. These TNTs mediate the transfer of RBs from infected to adjacent non-infected cells. TNTs play a crucial role in the dissemination of Chlamydia, enabling pathogens to move across distances of up to 50 µm in size. C. trachomatis can spread directly from one cell to another through TNTs, even when the extracellular entry into host cells is inhibited (30). TNT-associated proteins play essential roles in RB cell division, highlighting the complexity of the obligate intracellular replication strategy of Chlamydia (Figure 2).
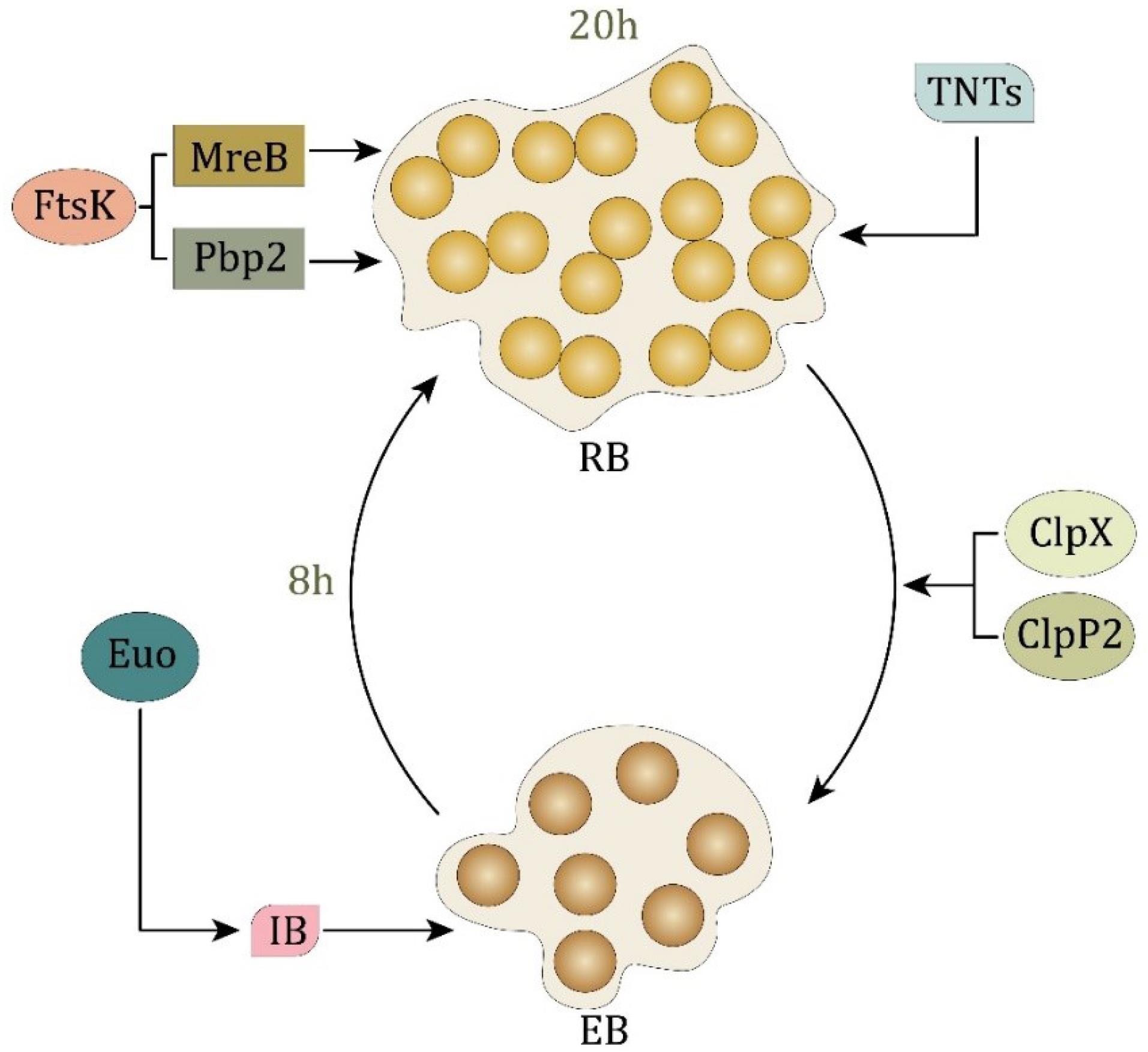
Figure 2.
The Biphasic Life Cycle of Chlamydia Alternating Between EB and RB. Note. EB: Elementary body; RB: Reticulate body; TNT: Tunnel nanotubes. EB represents the infectious, metabolically inactive form that transits into the replicative RB form approximately 8 hours post-infection. This transformation is regulated by Euo and inhibited by IB. RBs undergo proliferation and are subsequently reconverted into EBs, a process influenced by FtsK, MreB, and Pbp2 approximately 20 hours post-infection. Proteins ClpX and ClpP2 facilitate RB maintenance, whereas TNTs contribute to intercellular communication and dissemination
.
The Biphasic Life Cycle of Chlamydia Alternating Between EB and RB. Note. EB: Elementary body; RB: Reticulate body; TNT: Tunnel nanotubes. EB represents the infectious, metabolically inactive form that transits into the replicative RB form approximately 8 hours post-infection. This transformation is regulated by Euo and inhibited by IB. RBs undergo proliferation and are subsequently reconverted into EBs, a process influenced by FtsK, MreB, and Pbp2 approximately 20 hours post-infection. Proteins ClpX and ClpP2 facilitate RB maintenance, whereas TNTs contribute to intercellular communication and dissemination
Both C. pneumoniae and C. trachomatis, which are implicated in chronic chlamydial infections, are associated with high-morbidity diseases, particularly respiratory infections and reactive arthritis (31). A key contributor to chronic chlamydial infections is the metabolic activity of the host, which modulates chlamydial metabolism. This metabolic alteration can lead to persistent infections by inducing specific adaptive changes that enhance bacterial survival and proliferation (32,33). Additionally, the metabolic activity of the bacterium in its EB form contributes to the maintenance of its infectious potential and prolonged persistence in the host, both of which play a role in chronic chlamydial infections (34). Systemic C. trachomatis infection has been associated with systemic diseases, including asthma, Alzheimer’s disease, and reactive arthritis (35). The role of ClpX and ClpP2 in Chlamydia development highlights their potential as therapeutic targets. However, further studies are necessary to elucidate the precise mechanism of action.
Various Factors in the Increase in Chlamydia Pathogenicity
The most important pathogenicity factors of C. trachomatis are categorized based on their roles in bacterial survival, reproduction, and interaction with the host (Table 1 and Figure 3). The stages of the life cycle involve essential genes that regulate bacterial adaptation within the host cells. The regulatory elements of gene expression control pathogenicity by modulating important molecular signaling pathways. The type III secretion system is an important virulence determinant that enables bacterial effectors to manipulate host cellular functions, suppress immune responses, and facilitate intracellular survival. Specific virulence factors, including sctQ and copN, contribute to bacterial replication and host cell modulation.
Table 1.
Pathogenicity Factors of Chlamydia trachomatis and Their Roles
|
Specific Pathogenicity Factors
|
Details
|
Reference(s)
|
| Life cycle stages |
A significant portion of C. trachomatis pathogenicity is attributed to essential genes for each life cycle stage. The survival and proliferation of bacteria within host cells depend on these genes. |
(36-43) |
| Regulatory elements of gene expression |
The pathogenicity of this bacterium is influenced by several factors. Understanding the regulation of gene expression is crucial for understanding molecular processes underlying pathogenicity. |
| T3SS |
The most significant pathogenic factors in C. trachomatis are the secretion system and the variables affecting it. These elements modify the host cell functions to the advantage of the bacterium. Through T3SS, bacteria inject effector proteins into target cells, altering host signaling pathways and inhibiting immune responses. These components are crucial for the pathogenesis of infection. |
| Specific virulence factors |
Certain C. trachomatis factors significantly influence the pathogenicity. These components interact with host cells to promote microbial survival and replication. Various genes affect these classifications and have distinct functions. For example, sctQ genes play distinct roles in T3SS copN |
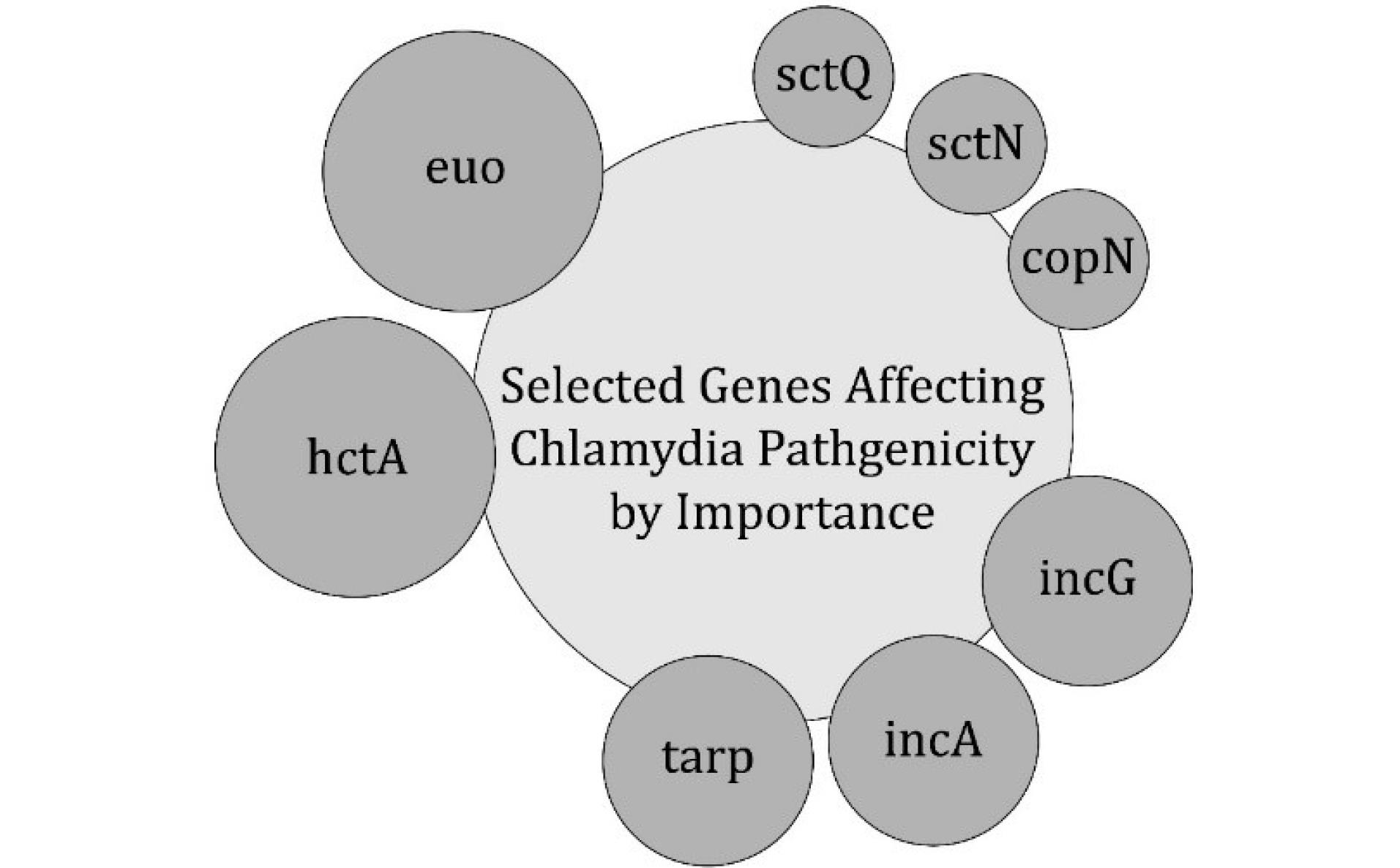
Figure 3.
Selected Genes Influencing the Pathogenicity of Chlamydia trachomatis, Categorized According to Their Relative Importance. Note. T3SS: Type III secretion system; C. trachomatis: Chlamydia trachomatis. Genes such as euo and hctA regulate bacterial development and the transition between different stages of the life cycle. Tarp plays a crucial role in host cell invasion. Sct, sctN, and copN are components of the T3SS, which facilitates bacterial manipulation of host cell functions. The inclusion of membrane proteins (IncA and IncG) contributes to the intracellular survival and evasion of the immune system. This schematic diagram illustrates the key molecular determinants of C. trachomatis pathogenesis
.
Selected Genes Influencing the Pathogenicity of Chlamydia trachomatis, Categorized According to Their Relative Importance. Note. T3SS: Type III secretion system; C. trachomatis: Chlamydia trachomatis. Genes such as euo and hctA regulate bacterial development and the transition between different stages of the life cycle. Tarp plays a crucial role in host cell invasion. Sct, sctN, and copN are components of the T3SS, which facilitates bacterial manipulation of host cell functions. The inclusion of membrane proteins (IncA and IncG) contributes to the intracellular survival and evasion of the immune system. This schematic diagram illustrates the key molecular determinants of C. trachomatis pathogenesis
Diagnosis and Intervention of Chlamydia Infection
Multiple Methods for the Diagnosis of an Infection Caused by Chlamydia
Conventional Techniques
C. trachomatis can be diagnosed through conventional diagnostic methods, such as cytological analysis, antigen detection, cell culture, and serological assays (Figure 4). Nucleic acid amplification tests are the most sensitive, whereas cell culture is the gold standard for specificity (44).
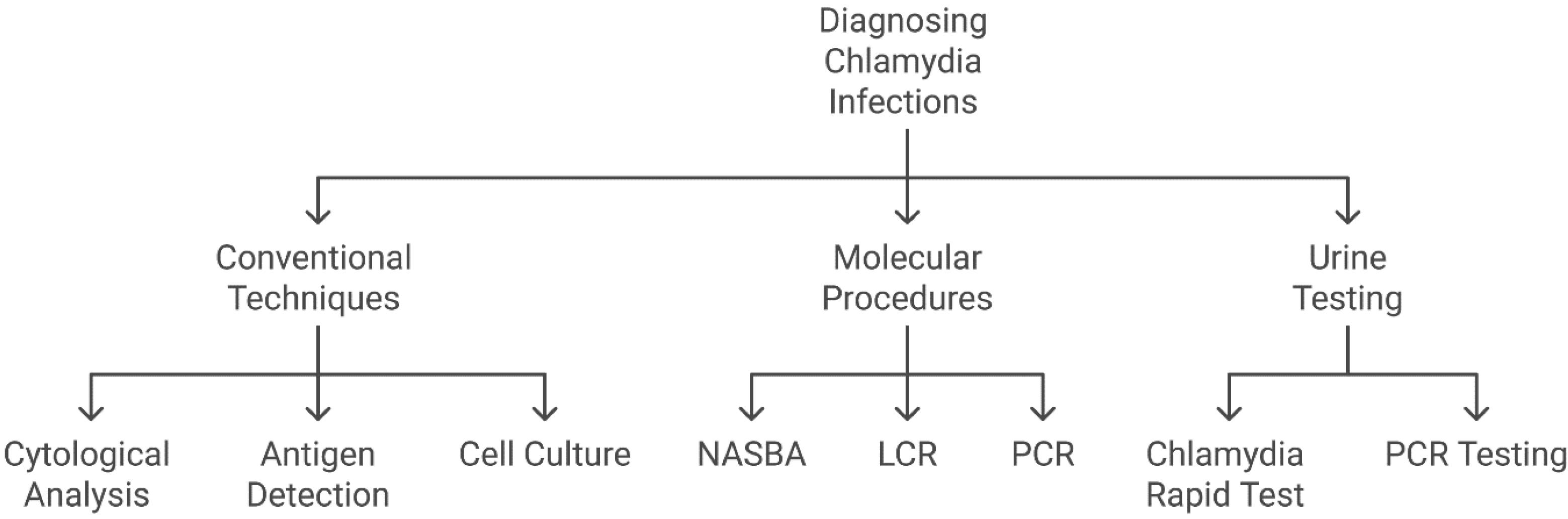
Figure 4.
Overview of Diagnostic Techniques for Chlamydia trachomatis Infection. Note. PCR: Polymerase chain reaction; LCR: Ligase chain reaction; NASBA: Nucleic acid sequence-based amplification.These methods are categorized into three primary groups, including conventional techniques (e.g., cytological analysis, antigen detection, and cell culture, which have been traditionally employed but frequently demonstrate lower sensitivity), molecular procedures (e.g., PCR, LCR, and NASBA, which offer high sensitivity and specificity through the detection of nucleic acids), and urine testing (comprising PCR-based methods and rapid tests, providing non-invasive, efficient, and user-friendly diagnostic options).
.
Overview of Diagnostic Techniques for Chlamydia trachomatis Infection. Note. PCR: Polymerase chain reaction; LCR: Ligase chain reaction; NASBA: Nucleic acid sequence-based amplification.These methods are categorized into three primary groups, including conventional techniques (e.g., cytological analysis, antigen detection, and cell culture, which have been traditionally employed but frequently demonstrate lower sensitivity), molecular procedures (e.g., PCR, LCR, and NASBA, which offer high sensitivity and specificity through the detection of nucleic acids), and urine testing (comprising PCR-based methods and rapid tests, providing non-invasive, efficient, and user-friendly diagnostic options).
Molecular Procedure
Chlamydia is detected, quantified, and identified through molecular diagnostic techniques, such as nucleic acid sequence-based amplification (NASBA), ligase chain reaction (LCR), and polymerase chain reaction (PCR). Compared to conventional PCR, real-time (RT)-PCR offers enhanced sensitivity in detecting genetic variations and is widely used for pathogen identification (45). These molecular techniques (Figure 4) are critical for the sensitive and accurate identification of Chlamydia infections. Advanced methods, such as PCR, LCR, NASBA, and RT-PCR, enable medical practitioners to identify and diagnose Chlamydia infections with high specificity and accuracy. Physicians can accurately detect Chlamydia infections using molecular diagnostic techniques, thereby facilitating early disease management and treatment. Moreover, the most accurate and advised technique for identifying C. trachomatis infections is the nucleic acid amplification test, particularly when the infection affects the lower genital tract, anus, throat, or ocular epithelium (46). High accuracy, sensitivity, and stability were achieved in the concurrent identification of three animal Chlamydia species, including C. psittaci, C. abortus, and C. pecorum, via multiplex RT TaqMan-MGB PCR processes (47).
Urine Testing for Identifying Chlamydia Infections
Several studies have reported the effectiveness of urine tests in detecting C. trachomatis infections (Figure 4). With an overall sensitivity of 94.6% and a relative accuracy of > 99.9%, the Chlamydia rapid test showed high accuracy in detecting bacteria in urine samples, yielding a total accuracy of 97.9%. These findings indicate that this technique offers a rapid and accurate approach to diagnosing sexually transmitted Chlamydia infections (48). Similarly, reactive arthritis (ReA) associated with C. trachomatis has been reliably detected in its early stages through PCR analysis of urine samples using specific primers; C. trachomatis DNA was identified in 36% of ReA patients (49).
Expansion of Effective Drug Therapies and Novel Treatment Approaches
This overview presented conventional, alternative, and novel therapeutic strategies for C. trachomatis infection (Table 1). Conventional antibiotics, such as doxycycline and azithromycin, primarily target bacterial proteins or DNA synthesis and are approved for clinical use. Alternative therapies, including plant extracts and host defense peptides, are being investigated for their antimicrobial properties. Novel therapeutic approaches, such as spider venom and ezrin-derived peptides, are currently in the research phase and have demonstrated potential for innovative treatment strategies. Table 2 summarizes the mechanisms of action, current status, and references for each category, providing insights into both the established and emerging treatment options.
Table 2.
Conventional, Alternative, and Novel Therapeutic Strategies for Chlamydia trachomatis Infection
|
Category
|
Medication/Agent
|
Mechanism of Action
|
Current Status
|
Reference(s)
|
| Conventional drugs |
Doxycycline |
Inhibits bacterial protein synthesis by binding to the 30S ribosomal subunit. |
Approved for use |
(50) |
| Azithromycin |
Binds to the 50S ribosomal subunit, preventing bacterial protein synthesis. |
Approved for use |
(51) |
| Ofloxacin |
Inhibits bacterial DNA gyrase, interfering with DNA replication. |
Approved for use |
(52) |
| Erythromycin |
Inhibits bacterial protein synthesis by binding to the 50S ribosomal subunit. |
Approved for use |
(50) |
| Trimethoprim-sulfamethoxazole |
Inhibits bacterial folic acid synthesis, essential for DNA and RNA production. |
Approved for use |
(50) |
| Alternative therapies |
Plant extracts |
Recent studies have identified bisabolane sesquiterpenes isolated from Ligularia narynensis, which demonstrated anti-chlamydial activity against C. abortus in host cells, suggesting that specific plant-derived compounds may serve as potential therapeutic candidates.
A comprehensive study assessed the antimicrobial properties of various plant extracts against C. trachomatis and identified Artemisia inculta as a potent inhibitor of Chlamydia infection in vitro. These findings highlight its potential as a promising candidate for alternative therapeutic strategies. |
Experimental |
(53,54) |
| Host defense peptides |
Mimics natural peptides such as LL-37, disrupting bacterial membranes and modulating immune responses. |
Experimental |
| Novel therapies |
Spider venom peptide |
Lycotoxin-Pa2a has antimicrobial properties and reduces inflammation; it has the potential for gene therapy. |
In the research phase |
(55) |
| (HDPs |
HDPs are capable of compromising the integrity of Chlamydia EB, thereby preventing their attachment and entry into host cells. Research indicates that peptides such as LL-37 and melittin possess potent anti-chlamydial properties, significantly lowering infection rates. Considering the constraints of existing antibiotic treatments, especially with the rise of resistance, HDPs offer a promising strategy for developing novel therapies against Chlamydia infections. |
In the research phase |
(56,57) |
| Ezrin peptides (human ezrin peptide one: HEP-1) |
Ezrin-derived peptides, particularly HEP-1, have been recognized as promising therapeutic strategies for managing Chlamydia infections. HEP-1, a 14-amino acid peptide, enhances adaptive immunity and has demonstrated effectiveness against various STIs, including Chlamydia, in clinical applications in Russia since 2001. Its mechanism of action involves stimulating immune responses while downregulating pro-inflammatory cytokines such as IL-6, which play a critical role in infection pathogenesis. |
In the research phase |
(58) |
| Limitations(s) |
Gene and peptide therapy |
These therapeutic strategies exhibit potential; however, their clinical translation necessitates rigorous validation of efficacy and safety. Moreover, gene therapy delivery remains challenging due to limitations in cellular uptake and the stability of therapeutic molecules. |
- |
(59,60) |
| Plant extracts |
The safety profiles of numerous plant-derived compounds remain inadequately characterized, with some demonstrating potential cytotoxicity even at therapeutically relevant concentrations. Furthermore, the lack of well-controlled clinical trials assessing the efficacy of these botanical extracts against Chlamydia trachomatis significantly hinders their integration into mainstream medical practice. |
- |
(61,62) |
Note. EB: Elementary body; IL: Interleukin; HDPs: Host protection peptides; STIs: sexually transmitted infections.
Challenges in Developing Gene- and Peptide-Based Treatments for Chlamydia
Several key challenges exist in the development of peptide and gene therapy for Chlamydia, including accurate antigen recognition and genetic constraints.
-
Accurate antigen recognition: Bioinformatic studies have identified unique antigen targets in the outer membrane proteins of C. trachomatis, suggesting a potential candidate for T-cell activation in Chlamydia treatment. These findings also provide insights into novel vaccine strategies aimed at preventing Chlamydia transmission (63).
-
Genetic constraints: C. trachomatis is challenging to genetically manipulate due to its complex and obligatory intracellular life cycle. Although significant progress has been made, including the use of suicide vectors and targeted mutagenesis techniques, several obstacles remain to be addressed (64,65).
The Increasing Threat of Drug Resistance to Chlamydia trachomatis Treatment
Although antibiotic resistance in Chlamydia has been reported, it does not appear to be widespread. Resistance to azithromycin in C. trachomatis has been rarely reported, although it has been observed in laboratory settings under specific conditions (66). Resistance to tetracyclines and macrolides remains uncommon; however, heterotypic resistance has been observed in certain cases, occurring under specific conditions and high bacterial loads (67). Growing concerns have emerged regarding treatment efficacy, as increasing antibiotic resistance to Mycoplasma hominis and C. trachomatis is increasing (68). In C. trachomatis, efflux pumps significantly contribute to antibiotic resistance by actively expelling antibiotics, thereby reducing their intracellular concentration and efficacy. Studies have identified compounds, such as selenocompounds, that inhibit these efflux pumps and increase antibiotic efficacy against C. trachomatis (69). Furthermore, certain isoflavones, notably biochanin A, have been shown to inhibit efflux pumps in C. trachomatis, thereby enhancing bacterial susceptibility to antimicrobial agents. This highlights the potential of efflux pump inhibitors as adjunctive treatments for resistance (70). Bacterial antibiotic resistance is largely mediated by the multiple transferable resistance (Mtr) efflux system. This system, designated MtrCDE, comprises three essential components. (1) MtrC is a periplasmic membrane fusion protein, and (2) MtrD is an inner membrane transporter. In addition, MtrE is an outer membrane channel. These elements function synergistically to form a complex that actively extrudes deleterious substances, thereby reducing the intracellular concentration of antimicrobial agents and facilitating drug resistance (71). Fluoroquinolone resistance typically involves mutations in gyrA, while macrolide resistance, particularly to azithromycin, is associated with mutations in the 23S rRNA gene. Additionally, mutations in rpoB, such as H471Y, are linked to rifampin resistance. Fosfomycin resistance arises due to modifications in the murA gene, altering the antibiotic’s binding site (72,73). The integration of exogenous genomic elements, such as the tet gene, into bacterial chromosomes is strongly linked to antibiotic resistance. This phenomenon has been observed in related species, such as Chlamydia suis, and can be transmitted to C. trachomatis through horizontal gene transfer (74). The spread of antibiotic resistance has been exacerbated by the excessive and inappropriate use of these agents in both clinical and agricultural settings, coupled with inadequate diagnostic methodologies. Furthermore, the practice of prophylactically administering antibiotics to asymptomatic individuals who have been in contact with infected persons has contributed to unnecessary exposure to these antimicrobial compounds. Implement targeted prescription protocols and “assess-then-postpone” strategies to minimize unnecessary antibiotic administration (75). The emergence of antibiotic resistance in STIs, including Chlamydia, may lead to increased healthcare costs and a higher disease burden, particularly in disadvantaged communities (76).
Future Prospects for Developing a Vaccine Against Chlamydia Infection
Recent advancements in C. trachomatis vaccine development have been observed. Table 3 elucidates the key research areas, including the successful completion of the first clinical trial, advancements in multi-epitope immunization strategies, and insights into immune response optimization. Animal model studies have demonstrated promising results, particularly for Chlamydia protease-like activity factor (CPAF)-targeted vaccines in porcine models. Furthermore, research on attenuated live vaccines suggests their potential as a viable strategy for long-term protection against Chlamydia infection. These findings contribute to ongoing efforts to develop an efficacious and durable Chlamydia vaccine.
Table 3.
Vaccine Development Against Chlamydia Infection
|
Research Area
|
Key Findings
|
Reference(s)
|
| First clinical trial |
The Phase 1 clinical trial of the CTH522 Chlamydia vaccine focused on investigating its safety and ability to trigger an immune response in humans. This randomized, double-blind, and placebo-controlled study evaluated the vaccine with two distinct adjuvants (CAF01 liposomes and aluminum hydroxide). The findings indicated that both formulations were well-tolerated and exhibited no significant safety concerns. However, the CAF01-adjuvanted vaccine demonstrated a stronger immunogenic profile compared to the aluminum hydroxide formulation. These results highlight the potential of the CTH522:CAF01 combination for further clinical advancement. The trial primarily assessed safety, while the secondary focus was on humoral immunity, measured by anti-CTH522 IgG seroconversion. The vaccine elicited a strong immune response, significantly increasing IgG levels. |
(77,78) |
| Multi-epitope immunization |
The exploration of multi-epitope vaccines, incorporating a diverse range of antigens from human papillomavirus, Chlamydia trachomatis, and herpes simplex, offers a novel approach to enhance both humoral and cellular immune responses, particularly for sexually transmitted diseases. |
(79) |
| Immune response optimization |
Immune response analysis indicates that effective vaccination requires the activation of a balanced Th1 and Th2 immune response. These findings suggest a pathway for the development of safer and more effective Chlamydia vaccines, aligning immune system activation with vaccine efficacy. |
(80) |
| Animal model research |
Animal model studies, particularly in porcine models, have revealed that vaccines targeting the CPAF protein, when combined with the TriAdj adjuvant, can induce robust immune responses, potentially accelerating the transition of vaccine candidates from preclinical to clinical stages. |
(81) |
| Attenuated live vaccines |
A genetically modified strain of Chlamydia muridarum has demonstrated enhanced protective capabilities against genital Chlamydia challenges, with a reduced ability to infect the genital tract, suggesting that live attenuated vaccines may serve as an effective strategy for preventing Chlamydia infections. |
(82) |
| Vaccine efficacy and challenges |
The initial trials of the CTH522 vaccine candidate have been completed, confirming its safety and immunogenicity in Phase I/II studies. However, further research is required to evaluate its efficacy in infection prevention. Preclinical studies on multi-epitope vaccines, designed using computational immunology approaches, have shown promise in eliciting strong immune responses in animal models. Live-attenuated vaccines demonstrate significant potential; nonetheless, they encounter substantial regulatory hurdles for approval. |
(83-86) |
Note. IgG: Immunoglobulin G; Th: T-helper; CPAF: Chlamydia protease-like activity factor.
Immune System Avoidance of Chlamydia
Evasion From Neutrophils and Dendritic Cells
Neutrophils and dendritic cells (DCs) are critical components of the early immune response that employ phagocytosis and antigen presentation to combat pathogens. However, C. trachomatis uses multiple mechanisms to evade these defense systems. The bacterium produces nucleases and proteases to degrade neutrophil extracellular traps and secretes CPAF to inhibit FPR2-mediated neutrophil recruitment (87-89). Furthermore, C. trachomatis modifies Rab proteins in DCs, which disturbs effective antigen presentation via major histocompatibility complex (MHC)-I and MHC-II pathways. Certain strains induce apoptosis in both neutrophils and DCs through the caspase 3/7 pathway, leading to impaired immune signaling and clearance (90-92).
Manipulation of Mast Cells, Eosinophils, and Natural Killer Cells
Mast cells and eosinophils play critical roles in allergic and inflammatory responses. C. trachomatis infection induces an excessive interleukin (IL)-4 response, resulting in host tissue damage and immune dysregulation (93-95). Further, the bacterium alters NK cell function by modulating MHC-I expression and evading NKG2D receptor-mediated cytotoxicity, allowing immune evasion (96,97).
Immune Evasion Strategies of Chlamydia in Macrophages
MQs are crucial for intracellular pathogen clearance through cytokine signaling and nitric oxide (NO) production. C. trachomatis modulates MQ metabolic pathways by inducing a shift toward glycolysis and regulating NO synthesis via inducible nitric oxide synthase and signal transducer and activator of transcription 1 pathways (98,99). Moreover, the bacterium promotes pyroptosis through gasdermin activation, leading to the excessive release of IL-1β and IL-18, which may contribute to persistent infection (100,101) (Figure 5).
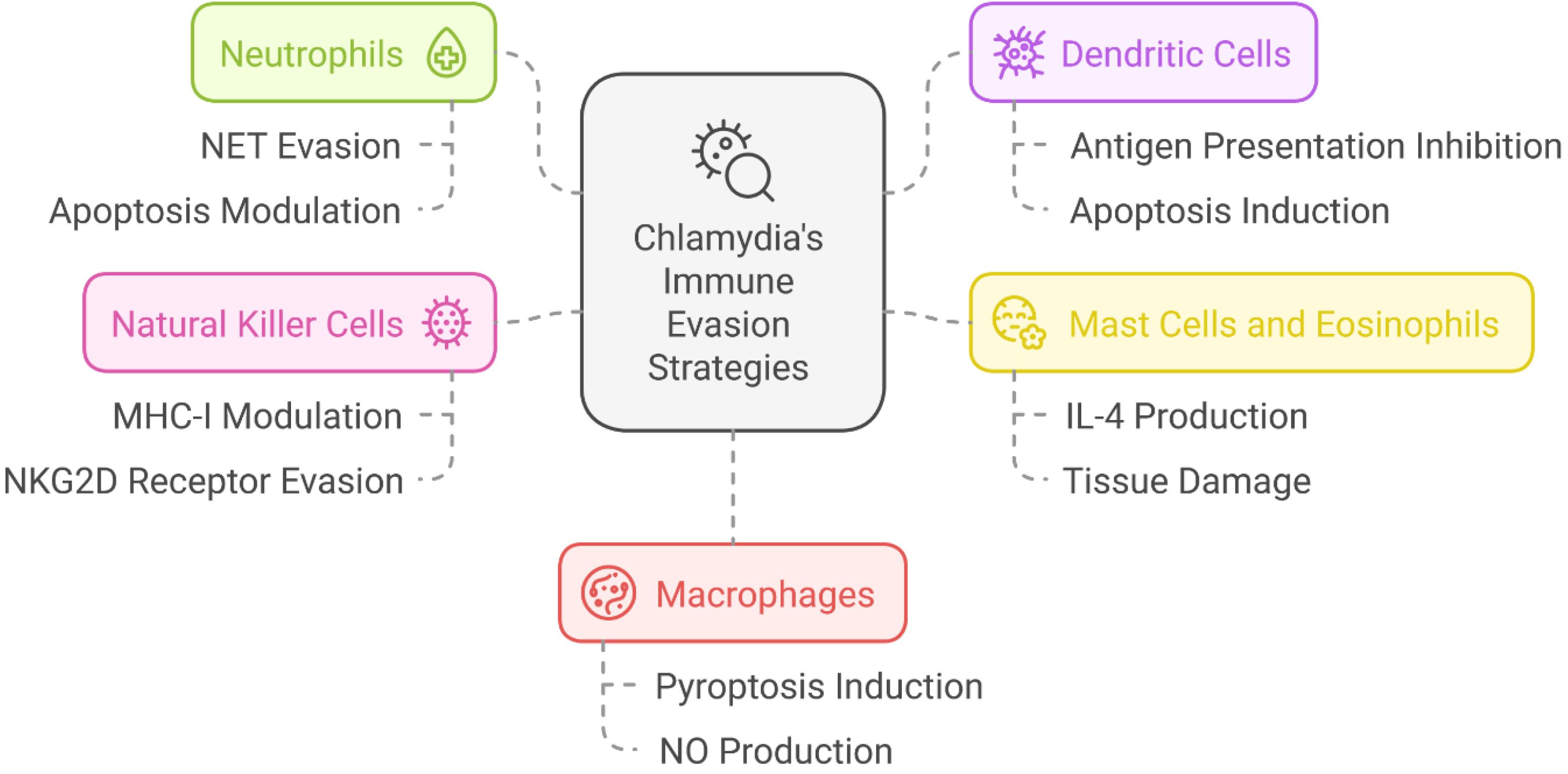
Figure 5.
Chlamydia trachomatis Employing Diverse Immune Evasion Strategies to Establish Persistent Infection. Note. NET: Neutrophil extracellular trap; NK: Natural killer; NO: Nitric oxide; MHC: Major histocompatibility complex; IL: Interleukin; C. trachomatis: Chlamydia trachomatis. These mechanisms involve the modulation of key immune cells: (1) neutrophils, by evading NETs and modulating apoptosis, (2) dendritic cells, by inhibiting antigen presentation and inducing apoptosis, (3) NK cells, by modulating MHC-I expression and evading NKG2D receptor-mediated recognition, (4) macrophages, by inducing pyroptosis and regulating NO production, and (5) mast cells and eosinophils, by stimulating IL-4 production and contributing to tissue damage. These strategies enable C. trachomatis to evade immune detection and establish chronic infections
.
Chlamydia trachomatis Employing Diverse Immune Evasion Strategies to Establish Persistent Infection. Note. NET: Neutrophil extracellular trap; NK: Natural killer; NO: Nitric oxide; MHC: Major histocompatibility complex; IL: Interleukin; C. trachomatis: Chlamydia trachomatis. These mechanisms involve the modulation of key immune cells: (1) neutrophils, by evading NETs and modulating apoptosis, (2) dendritic cells, by inhibiting antigen presentation and inducing apoptosis, (3) NK cells, by modulating MHC-I expression and evading NKG2D receptor-mediated recognition, (4) macrophages, by inducing pyroptosis and regulating NO production, and (5) mast cells and eosinophils, by stimulating IL-4 production and contributing to tissue damage. These strategies enable C. trachomatis to evade immune detection and establish chronic infections
Conclusion
Chlamydia, particularly C. trachomatis, is one of the most prevalent sexually transmitted bacterial pathogens. It primarily targets the epithelial cells of the human reproductive tract. Concerns associated with chlamydial infections include pelvic inflammatory disease, infertility, and trachoma. The public health and individual consequences of these complications are significant.
Therefore, effective strategies for the prevention and management of chlamydial infections are crucial. This study investigated the defense mechanisms employed by C. trachomatis to evade the human immune system and its replication cycle. Furthermore, this study explored the potential of novel antibiotic- and peptide-based therapies to address the emerging issue of antibiotic resistance in C. trachomatis. By understanding the intricate mechanisms of chlamydial pathogenesis, this study attempted to facilitate the development of more effective treatment options. This study also discussed how preventive medicine measures could help interrupt transmission dynamics to eradicate this infection. To develop more sophisticated treatments for Chlamydia infections, further research is needed on the development of vaccines targeting key virulence factors. This study underscores the importance of targeting virulence factors such as TNTs in developing next-generation therapies. Future research should focus on translating these findings into clinical implications.
Competing Interests
The authors declare that they have no conflict of interests.
Ethical Approval
This article is based on previously conducted studies and does not contain any new studies with human participants or animals performed by any of the authors.
Funding
No funding or sponsorship was received for this study or publication of this article.
References
- Tagini F, Greub G. [Chlamydial infections: epidemiology, pathogenesis, diagnosis and treatments]. Rev Med Suisse 2018;14(618):1620-5. [French].
- Misra SK, Pundir A. Chlamydia: the secret enemy from the past to present, and future. In: Sarier M, ed. Chlamydia-Secret Enemy from Past to Present. IntechOpen; 2023. doi: 10.5772/intechopen.110902.
- Cheong HC, Lee CY, Cheok YY, Tan GM, Looi CY, Wong WF. Chlamydiaceae: diseases in primary hosts and zoonosis. Microorganisms 2019; 7(5):148. doi: 10.3390/microorganisms7050146 [Crossref] [ Google Scholar]
- Gunn A, Lofstedt R. Chlamydiaceae: an update on nomenclature. Vet Rec 2016; 179(8):193-4. doi: 10.1136/vr.i4193 [Crossref] [ Google Scholar]
- Brunham RC, Rey-Ladino J. Immunology of Chlamydia infection: implications for a Chlamydia trachomatis vaccine. Nat Rev Immunol 2005; 5(2):149-61. doi: 10.1038/nri1551 [Crossref] [ Google Scholar]
- Albini S, Marti H, Imkamp F, Borel N. [Update on the zoonotic potential of Chlamydia]. Schweiz Arch Tierheilkd 2023; 165(3):165-78. doi: 10.17236/sat00387.[German] [Crossref] [ Google Scholar]
- Geisler WM. Duration of untreated, uncomplicated Chlamydia trachomatis genital infection and factors associated with Chlamydia resolution: a review of human studies. J Infect Dis 2010; 201 Suppl 2:S104-13. doi: 10.1086/652402 [Crossref] [ Google Scholar]
- Hocking JS, Huston WM, Chen M. Chlamydia trachomatis infection. In: Bachmann LH, ed. Sexually Transmitted Infections in HIV-Infected Adults and Special Populations: A Clinical Guide. Cham: Springer; 2017. p. 51-67. doi: 10.1007/978-3-319-56694-8_3.
- Daloglu H. Childhood Chlamydia infections. In: Sarier M, ed. Chlamydia-Secret Enemy from Past to Present. IntechOpen; 2023. doi: 10.5772/intechopen.111712.
- Piñeiro L, Galán JC, Vall-Mayans M. Infections caused by Chlamydia trachomatis (including lymphogranuloma venereum) and Mycoplasma genitalium. Enferm Infecc Microbiol Clin (Engl Ed) 2019; 37(8):525-34. doi: 10.1016/j.eimc.2019.01.014 [Crossref] [ Google Scholar]
- Hogan RJ, Mathews SA, Mukhopadhyay S, Summersgill JT, Timms P. Chlamydial persistence: beyond the biphasic paradigm. Infect Immun 2004; 72(4):1843-55. doi: 10.1128/iai.72.4.1843-1855.2004 [Crossref] [ Google Scholar]
- Hu VH, Holland MJ, Burton MJ. Trachoma: protective and pathogenic ocular immune responses to Chlamydia trachomatis. PLoS Negl Trop Dis 2013; 7(2):e2020. doi: 10.1371/journal.pntd.0002020 [Crossref] [ Google Scholar]
- Connell CM, Ferone ME. Chlamydia trachomatis genital infections. Microb Cell 2016; 3(9):390-403. doi: 10.15698/mic2016.09.525 [Crossref] [ Google Scholar]
- Koca Ö. The laboratory diagnosis of Chlamydia infections. In: Sarier M, ed. Chlamydia-Secret Enemy from Past to Present. IntechOpen; 2023. doi: 10.5772/intechopen.110464.
- Holló P, Jókai H, Herszényi K, Kárpáti S. [Genitourethral infections caused by D-K serotypes of Chlamydia trachomatis]. Orv Hetil 2015; 156(1):19-23. doi: 10.1556/oh.2015.30078.[Hungarian] [Crossref] [ Google Scholar]
- Ceovic R, Gulin SJ. Lymphogranuloma venereum: diagnostic and treatment challenges. Infect Drug Resist 2015; 8:39-47. doi: 10.2147/idr.S57540 [Crossref] [ Google Scholar]
- Brunham RC. Using epidemiology, immunology, and genomics to study the biology of Chlamydia trachomatis. Sex Transm Dis 2021; 48(5):319-22. doi: 10.1097/olq.0000000000001316 [Crossref] [ Google Scholar]
- Rabasseda X, Morré SA, Ouburg S. Bioinformatic approaches to the study of chlamydial diseases. Drugs Today (Barc) 2009; 45 Suppl B:173-87. [ Google Scholar]
- Poston TB. Advances in vaccine development for Chlamydia trachomatis. Pathog Dis 2024;82. doi: 10.1093/femspd/ftae017.
- Chiarelli TJ, Grieshaber NA, Appa C, Grieshaber SS. Computational modeling of the chlamydial developmental cycle reveals a potential role for asymmetric division. mSystems 2023; 8(2):e0005323. doi: 10.1128/msystems.00053-23 [Crossref] [ Google Scholar]
- Appa CR, Grieshaber NA, Yang H, Omsland A, McCormick S, Chiarelli TJ, et al. The chlamydial transcriptional regulator Euo is a key switch in cell form developmental progression but is not involved in the committed step to the formation of the infectious form. bioRxiv [Preprint]. May 18, 2024. Available from: https://www.biorxiv.org/content/10.1101/2024.05.17.594781v1.
- Wan FY. Growth of RB population in the conversion phase of Chlamydia life cycle. Commun Appl Math Comput 2024; 6(1):90-112. doi: 10.1007/s42967-022-00226-w [Crossref] [ Google Scholar]
- Abdelrahman YM, Belland RJ. The chlamydial developmental cycle. FEMS Microbiol Rev 2005; 29(5):949-59. doi: 10.1016/j.femsre.2005.03.002 [Crossref] [ Google Scholar]
- Nicholson TL, Olinger L, Chong K, Schoolnik G, Stephens RS. Global stage-specific gene regulation during the developmental cycle of Chlamydia trachomatis. J Bacteriol 2003; 185(10):3179-89. doi: 10.1128/jb.185.10.3179-3189.2003 [Crossref] [ Google Scholar]
- Moulder JW. Interaction of chlamydiae and host cells in vitro. Microbiol Rev 1991; 55(1):143-90. doi: 10.1128/mr.55.1.143-190.1991 [Crossref] [ Google Scholar]
- Brockett MR, Lee J, Cox JV, Liechti GW, Ouellette SP. Chlamydia trachomatis encodes a dynamic, ring-forming bactofilin critical for maintaining cell size and shape. bioRxiv [Preprint]. October 20, 2020. Available from: https://www.biorxiv.org/content/10.1101/2020.10.19.346502v1.
- Ouellette SP, Karimova G, Subtil A, Ladant D. Chlamydia co-opts the rod shape-determining proteins MreB and Pbp2 for cell division. Mol Microbiol 2012; 85(1):164-78. doi: 10.1111/j.1365-2958.2012.08100.x [Crossref] [ Google Scholar]
- Ouellette SP, Lee J, Cox JV. Division without binary fission: cell division in the FtsZ-less Chlamydia. J Bacteriol 2020; 202(17):e00252-20. doi: 10.1128/jb.00252-20 [Crossref] [ Google Scholar]
- Wood NA, Blocker AM, Seleem MA, Conda-Sheridan M, Fisher DJ, Ouellette SP. The ClpX and ClpP2 orthologs of Chlamydia trachomatis perform discrete and essential functions in organism growth and development. mBio 2020; 11(5):e02016-20. doi: 10.1128/mBio.02016-20 [Crossref] [ Google Scholar]
- Jahnke R, Matthiesen S, Zaeck LM, Finke S, Knittler MR. Chlamydia trachomatis cell-to-cell spread through tunneling nanotubes. Microbiol Spectr 2022; 10(6):e0281722. doi: 10.1128/spectrum.02817-22 [Crossref] [ Google Scholar]
- Leonard CA, Borel N. Chronic chlamydial diseases: from atherosclerosis to urogenital infections. Curr Clin Microbiol Rep 2014; 1(3):61-72. doi: 10.1007/s40588-014-0005-8 [Crossref] [ Google Scholar]
- Rother M, Teixeira da Costa AR, Zietlow R, Meyer TF, Rudel T. Modulation of host cell metabolism by Chlamydia trachomatis. Microbiol Spectr 2019;7(3):10.1128/microbiolspec.bai-0012-2019. doi: 10.1128/microbiolspec.BAI-0012-2019.
- Vollmuth N, Schlicker L, Guo Y, Hovhannisyan P, Janaki-Raman S, Kurmasheva N. c-Myc plays a key role in IFN-γ-induced persistence of Chlamydia trachomatis. Elife 2022; 11:e76721. doi: 10.7554/eLife.76721 [Crossref] [ Google Scholar]
- Grieshaber S, Grieshaber N, Yang H, Baxter B, Hackstadt T, Omsland A. Impact of active metabolism on Chlamydia trachomatis elementary body transcript profile and infectivity. J Bacteriol 2018; 200(14):e00065-18. doi: 10.1128/jb.00065-18 [Crossref] [ Google Scholar]
- Cheok YY, Lee CYQ, Cheong HC, Looi CY, Wong WF. Chronic inflammatory diseases at secondary sites ensuing urogenital or pulmonary Chlamydia infections. Microorganisms 2020; 8(1):127. doi: 10.3390/microorganisms8010127 [Crossref] [ Google Scholar]
- Jury B, Fleming C, Huston WM, Luu LD. Molecular pathogenesis of Chlamydia trachomatis. Front Cell Infect Microbiol 2023; 13:1281823. doi: 10.3389/fcimb.2023.1281823 [Crossref] [ Google Scholar]
- Wimmi S. Molecular Function and Regulation of the Bacterial Injectisome [dissertation]. Philipps-Universität Marburg; 2021. doi: 10.17192/z2021.0241.
- Rucks EA. Type III secretion in Chlamydia. Microbiol Mol Biol Rev 2023; 87(3):e0003423. doi: 10.1128/mmbr.00034-23 [Crossref] [ Google Scholar]
- Grieshaber NA, Appa C, Ward M, Grossman A, McCormik S, Grieshaber BS, et al. The T3SS structural and effector genes of Chlamydia trachomatis are expressed in distinct phenotypic cell forms. bioRxiv [Preprint]. April 26, 2024. Available from: https://www.biorxiv.org/content/10.1101/2024.04.25.591156v1.
- Herrera CM, McMahon E, Swaney DL, Sherry J, Pha K, Adams-Boone K. The Chlamydia trachomatis Inc Tri1 interacts with TRAF7 to displace native TRAF7 interacting partners. Microbiol Spectr 2024; 12(7):e0045324. doi: 10.1128/spectrum.00453-24 [Crossref] [ Google Scholar]
- Pha K, Mirrashidi K, Sherry J, Tran CJ, Herrera CM, McMahon E, et al. The Chlamydia effector IncE employs two short linear motifs to reprogram host vesicle trafficking. bioRxiv [Preprint]. April 24, 2024. Available from: https://www.biorxiv.org/content/10.1101/2024.04.23.590830v1.
- Poston TB, Girardi J, Polson AG, Bhardwaj A, Yount KS, Jaras Salas I. Viral-vectored boosting of OmcB- or CPAF-specific T-cell responses fail to enhance protection from Chlamydia muridarum in infection-immune mice and elicits a non-protective CD8-dominant response in naïve mice. Mucosal Immunol 2024; 17(5):1005-18. doi: 10.1016/j.mucimm.2024.06.012 [Crossref] [ Google Scholar]
- Grishin AV, Luyksaar SI, Kapotina LN, Kirsanov DD, Zayakin ES, Karyagina AS. Identification of chlamydial T3SS inhibitors through virtual screening against T3SS ATPase. Chem Biol Drug Des 2018; 91(3):717-27. doi: 10.1111/cbdd.13130 [Crossref] [ Google Scholar]
- Peng L, Chen JL, Wang D. Progress and perspectives in point of care testing for urogenital Chlamydia trachomatis infection: a review. Med Sci Monit 2020; 26:e920873. doi: 10.12659/msm.920873 [Crossref] [ Google Scholar]
- Konuk EY. Molecular approaches to the diagnosis of Chlamydia. In: Sarier M, ed. Chlamydia-Secret Enemy from Past to Present. IntechOpen; 2023. doi: 10.5772/intechopen.109746.
- Meyer T. Diagnostic procedures to detect Chlamydia trachomatis infections. Microorganisms 2016; 4(3):25. doi: 10.3390/microorganisms4030025 [Crossref] [ Google Scholar]
- Nie F, Gong Q, Yang J, Xi C, Wang Y, Wang G. Establishment of a multiplex real-time TaqMan-MGB polymerase chain reaction (PCR) method for the simultaneous detection of three animal Chlamydia species. Med Sci Monit 2019; 25:9369-76. doi: 10.12659/msm.918344 [Crossref] [ Google Scholar]
- Lei Z, Feng Y, Junzhe Z. Performance evaluation of the Chlamydia rapid test for qualitative detection of chlamydial antigens in clinical diagnosis. MEDS Clin Med 2024; 5(1):158-63. doi: 10.23977/medsc.2024.050124 [Crossref] [ Google Scholar]
- Sharma M, Sharma S, Sharma A, Sharma K. Chlamydia trachomatis associated reactive arthritis: a urinary PCR based study. Indian Dermatol Online J 2020; 11(1):21-4. doi: 10.4103/idoj.IDOJ_410_19 [Crossref] [ Google Scholar]
- Johannisson G, Sernryd A, Lycke E. Susceptibility of Chlamydia trachomatis to antibiotics in vitro and in vivo. Sex Transm Dis 1979; 6(2):50-7. doi: 10.1097/00007435-197904000-00002 [Crossref] [ Google Scholar]
- Khosropour CM, Dombrowski JC, Barbee LA, Manhart LE, Golden MR. Comparing azithromycin and doxycycline for the treatment of rectal chlamydial infection: a retrospective cohort study. Sex Transm Dis 2014; 41(2):79-85. doi: 10.1097/olq.0000000000000088 [Crossref] [ Google Scholar]
- Oriel JD. Use of quinolones in chlamydial infection. Rev Infect Dis 1989; 11 Suppl 5:S1273-6. doi: 10.1093/clinids/11.supplement_5.s1273 [Crossref] [ Google Scholar]
- Gao N, He YL, Qi HM, Yang HY, Li GL, Li ZC. Bisabolane sesquiterpenes with anti-chlamydial activity isolated from Ligularianarynensis. Int J Mol Sci 2025; 26(3):1388. doi: 10.3390/ijms26031388 [Crossref] [ Google Scholar]
- Hamarsheh O, Amro A, Al-Zeer MA. In vitro antibacterial activity of selected Palestinian medicinal plants against Chlamydia trachomatis. Microbiol Res 2021; 12(3):656-62. doi: 10.3390/microbiolres12030047 [Crossref] [ Google Scholar]
- Lazarev VN, Polina NF, Shkarupeta MM, Kostrjukova ES, Vassilevski AA, Kozlov SA. Spider venom peptides for gene therapy of Chlamydia infection. Antimicrob Agents Chemother 2011; 55(11):5367-9. doi: 10.1128/aac.00449-11 [Crossref] [ Google Scholar]
- Brown MA, Potroz MG, Teh SW, Cho NJ. Natural products for the treatment of Chlamydiaceae infections. Microorganisms 2016; 4(4):39. doi: 10.3390/microorganisms4040039 [Crossref] [ Google Scholar]
- Di Francesco A, Favaroni A, Donati M. Host defense peptides: general overview and an update on their activity against Chlamydia spp. Expert Rev Anti Infect Ther 2013; 11(11):1215-24. doi: 10.1586/14787210.2013.841450 [Crossref] [ Google Scholar]
- Holms RD. Pandemics of sexually transmitted infections (STIs): clinical use of ezrin peptide therapy in Russia to treat and prevent Candida, Chlamydia, Trichomonas vaginalis, syphilis, HPV and herpes (HSV-1 & 2). Microbiol Infect Dis 2023; 7(3):1-21. [ Google Scholar]
- Loughran S, McCrudden CM, McCarthy HO. Designer peptide delivery systems for gene therapy. Eur J Nanomed 2015; 7(2):85-96. doi: 10.1515/ejnm-2014-0037 [Crossref] [ Google Scholar]
- Qin J. Gene and peptide in the treatment of Chlamydia trachomatis infections: beginning of an era?. Theoretical and Natural Science 2023; 4(1):18-25. doi: 10.54254/2753-8818/4/20220504 [Crossref] [ Google Scholar]
- Vaou N, Stavropoulou E, Voidarou C, Tsigalou C, Bezirtzoglou E. Towards advances in medicinal plant antimicrobial activity: a review study on challenges and future perspectives. Microorganisms 2021; 9(10):2041. doi: 10.3390/microorganisms9102041 [Crossref] [ Google Scholar]
- Tshikalange TE, Mamba P, Adebayo SA. Antimicrobial, antioxidant and cytotoxicity studies of medicinal plants used in the treatment of sexually transmitted diseases. Int J Pharmacogn Phytochem Res 2016; 8(11):1895-1. [ Google Scholar]
- Zhang T, Li H, Lan X, Zhang C, Yang Z, Cao W. The bioinformatics analyses reveal novel antigen epitopes in major outer membrane protein of Chlamydia trachomatis. Indian J Med Microbiol 2017; 35(4):522-8. doi: 10.4103/ijmm.IJMM_17_251 [Crossref] [ Google Scholar]
- Wan W, Li D, Li D, Jiao J. Advances in genetic manipulation of Chlamydia trachomatis. Front Immunol 2023; 14:1209879. doi: 10.3389/fimmu.2023.1209879 [Crossref] [ Google Scholar]
- Mueller KE, Wolf K, Fields KA. Gene deletion by fluorescence-reported allelic exchange mutagenesis in Chlamydia trachomatis. mBio 2016; 7(1):e01817-15. doi: 10.1128/mBio.01817-15 [Crossref] [ Google Scholar]
- Binet R, Bowlin AK, Maurelli AT, Rank RG. Impact of azithromycin resistance mutations on the virulence and fitness of Chlamydia caviae in guinea pigs. Antimicrob Agents Chemother 2010; 54(3):1094-101. doi: 10.1128/aac.01321-09 [Crossref] [ Google Scholar]
- Horner P. The case for further treatment studies of uncomplicated genital Chlamydia trachomatis infection. Sex Transm Infect 2006; 82(4):340-3. doi: 10.1136/sti.2005.019158 [Crossref] [ Google Scholar]
- Krupp K, Madhivanan P. Antibiotic resistance in prevalent bacterial and protozoan sexually transmitted infections. Indian J Sex Transm Dis AIDS 2015; 36(1):3-8. doi: 10.4103/0253-7184.156680 [Crossref] [ Google Scholar]
- Mosolygó T, Kincses A, Csonka A, Tönki Á S, Witek K, Sanmartín C. Selenocompounds as novel antibacterial agents and bacterial efflux pump inhibitors. Molecules 2019; 24(8):1487. doi: 10.3390/molecules24081487 [Crossref] [ Google Scholar]
- Hanski L, Genina N, Uvell H, Malinovskaja K, Gylfe Å, Laaksonen T. Inhibitory activity of the isoflavone biochanin A on intracellular bacteria of genus Chlamydia and initial development of a buccal formulation. PLoS One 2014; 9(12):e115115. doi: 10.1371/journal.pone.0115115 [Crossref] [ Google Scholar]
- Fairweather SJ, Gupta V, Chitsaz M, Booth L, Brown MH, O’Mara ML. Coordination of substrate binding and protonation in the N gonorrhoeae MtrD efflux pump controls the functionally rotating transport mechanism. ACS Infect Dis 2021; 7(6):1833-47. doi: 10.1021/acsinfecdis.1c00149 [Crossref] [ Google Scholar]
- Mestrovic T, Ljubin-Sternak S. Molecular mechanisms of Chlamydia trachomatis resistance to antimicrobial drugs. Front Biosci (Landmark Ed) 2018; 23(4):656-70. doi: 10.2741/4611 [Crossref] [ Google Scholar]
- Benamri I, Azzouzi M, Sanak K, Moussa A, Radouani F. An overview of genes and mutations associated with chlamydiae species’ resistance to antibiotics. Ann Clin Microbiol Antimicrob 2021; 20(1):59. doi: 10.1186/s12941-021-00465-4 [Crossref] [ Google Scholar]
- Suchland RJ, Sandoz KM, Jeffrey BM, Stamm WE, Rockey DD. Horizontal transfer of tetracycline resistance among Chlamydia spp in vitro. Antimicrob Agents Chemother 2009; 53(11):4604-11. doi: 10.1128/aac.00477-09 [Crossref] [ Google Scholar]
- Ahmed Z, Kent J, Bailey A. P18 Retrospective study of Chlamydia contacts and management. JAC Antimicrob Resist 2025;7(Suppl 1):dlae217.022. doi: 10.1093/jacamr/dlae217.022.
- Hussein S, Ahmed SK, Mohammed SM, Qurbani K, Ali S, Saber AF, et al. Recent developments in antibiotic resistance: an increasing threat to public health. Ann Anim Sci. 2024. doi: 10.2478/aoas-2024-0111.
- Abraham S, Juel HB, Bang P, Cheeseman HM, Dohn RB, Cole T. Safety and immunogenicity of the Chlamydia vaccine candidate CTH522 adjuvanted with CAF01 liposomes or aluminium hydroxide: a first-in-human, randomised, double-blind, placebo-controlled, phase 1 trial. Lancet Infect Dis 2019; 19(10):1091-100. doi: 10.1016/s1473-3099(19)30279-8 [Crossref] [ Google Scholar]
- Pollock KM, Borges ÁH, Cheeseman HM, Rosenkrands I, Schmidt KL, Søndergaard RE. An investigation of trachoma vaccine regimens by the Chlamydia vaccine CTH522 administered with cationic liposomes in healthy adults (CHLM-02): a phase 1, double-blind trial. Lancet Infect Dis 2024; 24(8):829-44. doi: 10.1016/s1473-3099(24)00147-6 [Crossref] [ Google Scholar]
- Dorosti H, Eskandari S, Zarei M, Nezafat N, Ghasemi Y. Design of a multi-epitope protein vaccine against herpes simplex virus, human papillomavirus and Chlamydia trachomatis as the main causes of sexually transmitted diseases. Infect Genet Evol 2021; 96:105136. doi: 10.1016/j.meegid.2021.105136 [Crossref] [ Google Scholar]
- Quigley BL, Timms P. The koala immune response to chlamydial infection and vaccine development-advancing our immunological understanding. Animals (Basel) 2021; 11(2):380. doi: 10.3390/ani11020380 [Crossref] [ Google Scholar]
- Proctor J, Stadler M, Cortes LM, Brodsky D, Poisson L, Gerdts V. A TriAdj-adjuvanted Chlamydia trachomatis CPAF protein vaccine is highly immunogenic in pigs. Vaccines (Basel) 2024; 12(4):423. doi: 10.3390/vaccines12040423 [Crossref] [ Google Scholar]
- Morrison SG, Giebel AM, Toh E, Banerjee A, Nelson DE, Morrison RP. A genital infection-attenuated Chlamydia muridarum mutant infects the gastrointestinal tract and protects against genital tract challenge. mBio 2020; 11(6):e02770-20. doi: 10.1128/mBio.02770-20 [Crossref] [ Google Scholar]
- Borges ÁH, Follmann F, Dietrich J. Chlamydia trachomatis vaccine development - a view on the current challenges and how to move forward. Expert Rev Vaccines 2022; 21(11):1555-67. doi: 10.1080/14760584.2022.2117694 [Crossref] [ Google Scholar]
- Tanner T, Medhavi FN, Richardson S, Omosun YO, Eko FO. In silico design and analysis of a multiepitope vaccine against Chlamydia. Pathog Dis 2024; 82:ftae015. doi: 10.1093/femspd/ftae015 [Crossref] [ Google Scholar]
- Aslam S, Ahmad S, Noor F, Ashfaq UA, Shahid F, Rehman A. Designing a multi-epitope vaccine against Chlamydia trachomatis by employing integrated core proteomics, immuno-informatics and in silico approaches. Biology (Basel) 2021; 10(10):997. doi: 10.3390/biology10100997 [Crossref] [ Google Scholar]
- Hafner LM, Timms P. Development of a vaccine for Chlamydia trachomatis: challenges and current progress. Vaccine (Auckl) 2015; 5:45-58. doi: 10.2147/vdt.s69487 [Crossref] [ Google Scholar]
- Kobayashi SD, Malachowa N, DeLeo FR. Neutrophils and bacterial immune evasion. J Innate Immun 2018; 10(5-6):432-41. doi: 10.1159/000487756 [Crossref] [ Google Scholar]
- Kobayashi SD, DeLeo FR, Quinn MT. Microbes and the fate of neutrophils. Immunol Rev 2023; 314(1):210-28. doi: 10.1111/imr.13163 [Crossref] [ Google Scholar]
- Rajeeve K, Das S, Prusty BK, Rudel T. Chlamydia trachomatis paralyses neutrophils to evade the host innate immune response. Nat Microbiol 2018; 3(7):824-35. doi: 10.1038/s41564-018-0182-y [Crossref] [ Google Scholar]
- Virok DP, Tömösi F, Keller-Pintér A, Szabó K, Bogdanov A, Poliska S. Indoleamine 2,3-dioxygenase cannot inhibit Chlamydia trachomatis growth in HL-60 human neutrophil granulocytes. Front Immunol 2021; 12:717311. doi: 10.3389/fimmu.2021.717311 [Crossref] [ Google Scholar]
- Vasilevsky S, Greub G, Nardelli-Haefliger D, Baud D. Genital Chlamydia trachomatis: understanding the roles of innate and adaptive immunity in vaccine research. Clin Microbiol Rev 2014; 27(2):346-70. doi: 10.1128/cmr.00105-13 [Crossref] [ Google Scholar]
- Bilenki L, Wang S, Yang J, Fan Y, Jiao L, Joyee AG. Adoptive transfer of CD8alpha + dendritic cells (DC) isolated from mice infected with Chlamydia muridarum are more potent in inducing protective immunity than CD8alpha- DC. J Immunol 2006; 177(10):7067-75. doi: 10.4049/jimmunol.177.10.7067 [Crossref] [ Google Scholar]
- Vicetti Miguel RD, Quispe Calla NE, Dixon D, Foster RA, Gambotto A, Pavelko SD. IL-4-secreting eosinophils promote endometrial stromal cell proliferation and prevent Chlamydia-induced upper genital tract damage. Proc Natl Acad Sci U S A 2017; 114(33):E6892-901. doi: 10.1073/pnas.1621253114 [Crossref] [ Google Scholar]
- Poli-Neto OB, Carlos D, Junior AF, Rosa-e-Silva JC, Meola J, Tiezzi D. Eutopic endometrium from women with endometriosis and chlamydial endometritis share immunological cell types and DNA repair imbalance: a transcriptome meta-analytical perspective. J Reprod Immunol 2021; 145:103307. doi: 10.1016/j.jri.2021.103307 [Crossref] [ Google Scholar]
- Chiba N, Shimada K, Chen S, Jones HD, Alsabeh R, Slepenkin AV. Mast cells play an important role in Chlamydia pneumoniae lung infection by facilitating immune cell recruitment into the airway. J Immunol 2015; 194(8):3840-51. doi: 10.4049/jimmunol.1402685 [Crossref] [ Google Scholar]
- Zajonc DM. Unconventional peptide presentation by classical MHC class I and implications for T and NK cell activation. Int J Mol Sci 2020; 21(20):7561. doi: 10.3390/ijms21207561 [Crossref] [ Google Scholar]
- Ibana JA, Aiyar A, Quayle AJ, Schust DJ. Modulation of MICA on the surface of Chlamydia trachomatis-infected endocervical epithelial cells promotes NK cell-mediated killing. FEMS Immunol Med Microbiol 2012; 65(1):32-42. doi: 10.1111/j.1574-695X.2012.00930.x [Crossref] [ Google Scholar]
- Min S, He P, Zhou Q, Chen H. The dual role of cytokine responses to Chlamydia trachomatis infection in host pathogen crosstalk. Microb Pathog 2022; 173(Pt A):105812. doi: 10.1016/j.micpath.2022.105812 [Crossref] [ Google Scholar]
- Wang F, Zhang S, Jeon R, Vuckovic I, Jiang X, Lerman A. Interferon gamma induces reversible metabolic reprogramming of M1 macrophages to sustain cell viability and pro-inflammatory activity. EBioMedicine 2018; 30:303-16. doi: 10.1016/j.ebiom.2018.02.009 [Crossref] [ Google Scholar]
- Wang Y, Gao W, Shi X, Ding J, Liu W, He H. Chemotherapy drugs induce pyroptosis through caspase-3 cleavage of a gasdermin. Nature 2017; 547(7661):99-103. doi: 10.1038/nature22393 [Crossref] [ Google Scholar]
- Yang S, Zeng J, Hao W, Sun R, Tuo Y, Tan L. IL-21/IL-21R promotes the pro-inflammatory effects of macrophages during C muridarum respiratory infection. Int J Mol Sci 2023; 24(16):12557. doi: 10.3390/ijms241612557 [Crossref] [ Google Scholar]